
There has been quite a bit of debate over the years in regards to whether or not virology adheres to the scientific method, with much of this debate focused on the lack of proper controls. The argument has centered on whether the controls that are sometimes used yet rarely described, known as mock infections, are even a valid control to begin with. For those who are unfamiliar, scientific controls are a check and balance system that are utilized during experimentation when researchers are attempting to determine the cause of an effect. Controls are designed to ensure that the presumed cause, known as the independent variable, is the only thing that could be causing the observed effect, known as the dependent variable:
“A study with control(s) is designed to ensure that the effects are due to the independent variables in the experiment. The use of controls allows to study one variable or factor at a time. It is, however, important that both the control and other (experimental) group(s) are exposed to the same conditions apart from the one variable under study. Doing so will help draw conclusions that are more accurate and reliable.”
https://www.biologyonline.com/dictionary/control
One of the main reasons I have not really focused as much on the lack of valid controls in virology is that there are other far more pressing issues which need to be addressed first before even getting to the experimental stage in order to discuss proper controls. Take a look at the steps of the scientific method for a moment:
- Observe a phenomenon
- Alternate hypothesis
- Independent variable (the presumed cause)
- Dependent variable (the observed effect)
- Control variables
- Null hypothesis
- Test/experiment
- Analyze the observation/data
- Validate/invalidate hypothesis
Determining the IV, DV, and controls come after observing a natural phenomenon and are established when creating a hypothesis. Virologists have a difficult time completing the first steps of the scientific method as, beyond finding people with similar symptoms, they are unable to observe how this natural phenomenon occurs. They are unable to see the assumed “virus” particles in nature. They can not witness these particles entering a human being and causing illness, nor can they see these particles being transferred from person-to-person and causing disease. Without the observation of how this natural phenomenon occurs, there is no ability to establish a valid hypothesis in order to design an experiment in order to test for the cause of an observed effect. This means that step number 2 of the scientific method is off the table. With the inability to successfully satisfy the first two steps of the scientific method, so too goes the ability to design and perform a scientific experiment. Thus, the argument in regards to whether or not the controls used during pseudoscientific cell culture experiments are valid or not is too far ahead of the game. The entire premise behind the cell culture experiments themselves is invalid, as was shown by the man who created the method.
The Establishment of the Cell Culture
Up to 1952, the virologists believed that a virus was a toxic protein or enzyme directly poisoning the body, and that it was somehow multiplied by the body itself and would spread
-Dr. Stefan Lanka https://viroliegyhome.files.wordpress.com/2022/08/wissenschafftplus-the-virus-misconception-part-1.pdf
in the body as well as between people and between animals. Medicine and science gave up on this idea in 1951, because the suspected virus had never been seen in an electron microscope and, above all, no control experiments had ever been carried out. It was acknowledged that even healthy animals, organs and tissue would release the same decay products during the decomposing process that had been previously misinterpreted as “viruses”. Virology had refuted itself.”
In the early 1950’s, virology was on its deathbed after decades of failing miserably in the attempts to purify and isolate the assumed “viral” particles in order to directly prove the existence and pathogenicity of these invisible entities. In all of the years of experimentation, virologists had nothing to show but indirect evidence of decay from human and animal tissue culture experiments claimed to be caused by the “virus” in question.

However, the experimental results were not specific to the presence of any assumed “virus” as the same state of decay was witnessed in tissues from healthy hosts. As Dr. Stefan Lanka pointed out in the above quote, virology had refuted its own pseudoscientific tissue culture experiments. On top of this, the findings of effects associated with a specific “virus” from one group of researchers could not be reproduced nor replicated by other groups of researchers. In fact, results were often contradictory towards what was considered established evidence. Researchers could not even agree on what exactly a “virus” was nor whether the experimental evidence offered was in fact valid or not. A great summary of this rift was presented in a 1999 essay by Karlheinz Lüdtke:
On the history of early virus research.
“With the “filterable” virus, something had been discovered which, according to the traditional concepts, which after all had mostly proved their worth in research into infectious diseases, could not be described in a way that all researchers could have shared. Very different interpretations of the nature of this phenomenon arose, which were put forward against each other. No experimental evidence for this or that concept, which all researchers should have accepted, could be presented by any side. In other words, the decision as to whether this or that explanation most accurately expresses the “true” nature of the virus could not be “objectified” empirically. Every version of the interpretation of the phenomenon remained open to attack, facts presented to the expert public could often be reinterpreted into fictions by opponents, who brought into play the dependence of the findings on the conditions of observation, the local situation of the experiments, the research-related nature of the attributions of characteristics, etc. as sources of error. For example, findings often reported by certain virus researchers at the time were not confirmed by other researchers as a result of their own experiments, or the observations could not be reproduced by all scientists working with the virus. Often, findings to the contrary were reported, or the findings that had been examined were considered artefacts. As with justification, reasons of various kinds could be invoked to reject the positions debated. Findings that were used to empirically confirm a suspected connection were often soon joined by negative findings reported by other researchers. However carefully and deliberately the techniques used in the experiments were employed, and despite the fact that each party could offer credible reasons for defending their respective positions and provide empirical evidence – which explains why “the various opponents ‘constructed’ widely diverging research objects which they identified as the ‘virus'” (van Helvoort 1994a: 202) – at no time did they offer compelling reasons that would have led the other party to finally abandon artifact accusations.”
While things were obviously not going well for the field, virology got a booster in the form of a new method purporting to establish the existence of these invisible entities. This was the cell culture method which was introduced in 1954 by John Franklin Enders during his attempts to identify a measles “virus.” As virologists could still not properly purify nor isolate the assumed “virus” particles directly from the fluids of a sick host, it was decided that the particles had to be grown in a culture instead as it was claimed that there were not enough particles present within the fluids. The “virus,” which could not be found directly inside the fluids in order to be studied properly, was somehow determined to need a host cell in order to replicate itself so that it could then be found and studied. The cell culture method was said to produce supposedly superior indirect evidence to that which had come before and it ultimately ended up reviving the dying field. To achieve this superior indirect evidence, Enders, a virologist who was awarded a Nobel Prize (also in 1954) for the discovery of the ability of poliomyelitis “viruses” to grow in various types of tissue cultures, replaced animal and human tissue cultures with animal and human cell cultures. In other words, Enders was ironically recognized with the Nobel Prize for the evidence he had gathered using the old refuted tissue culture practices which were subsequently replaced by his new cell culture method within the very same year.
When Enders created his cell culture method, it must be noted that steps one and two of the scientific method still had yet to be achieved, thus he was jumping into experimentation without observing a natural phenomenon, identifying the dependent variable (i.e. the effect), nor isolating the independent variable (i.e. “virus”) in order to establish a hypothesis to test against. Yet this did not stop his attempt to create pseudoscientific experiments designed to convince the world that these fictional entities exist. While I have covered the invalid cell culture practice in great detail here, I will provide a quick explanation for what Enders’ method consisted of. In his seminal measles paper, Enders took throat washings from suspected measles patients (which were obtained in gargled fat-free milk) and added the samples to human and monkey kidney cells. He mixed into the culture bovine amniotic fluid, beef embryo extract, horse serum, antibiotics, soybean trypsin inhibitor, and phenol red as an indicator of cell metabolism. This mixture was then incubated for days and the fluids were passaged on the 4th and 16th days. Enders eventually observed what is called the cytopathogenic effect, which is a pattern of damage appearing in the culture as the cell breaks apart and dies. This effect was assumed by Enders to be the direct result of the invisible “virus” within the fat-free milk throat washings as it breaks into the cell through lysis of the cell wall membrane which resulted in the death of the cell and the replication of the “virus.” In other words, he assumed that the cellular debris from a poisoned cell was not the broken pieces of a once intact cell but were instead the newly created “viral” copies. Despite the unscientific nature of the method, the cell culture was quickly established as the “gold standard” for “virus isolation” and is still used by virologists today:
Cell culture provides the optimum setting for the detection and identification of numerous pathogens of humans, which is achieved via virus isolation in the cell culture as the “gold standard” for virus discovery.
-WHO https://www.google.com/url?sa=t&source=web&rct=j&url=https://apps.who.int/iris/rest/bitstreams/1088173/retrieve&ved=2ahUKEwiQstb8yOj7AhWOIzQIHeqcDKAQFnoECB4QAQ&usg=AOvVaw2RBaX3pO2PZ3JyW0s7U3Ol
What needs to be understood is that, along with not having a valid independent variable in purified and isolated “viral” particles, Enders created his own dependent variable in the cytopathogenic effect. This effect is not a naturally observed phenomenon. It is only observed through experimentation and manipulation in the lab. Enders already assumed that the “virus” existed and that he would observe an effect if he added a sample containing the assumed “virus” to a cell culture. Once he witnessed this effect, Enders claimed that this was a direct result of the presence of a “virus” even though he could not see the “virus” within the cell culture. By doing so, Enders engaged in what is known as an affirming the consequent logical fallacy, normally expressed as such:

This is also an example of begging the question and circular reasoning as beautifully illustrated in this graph by Alec Zeck and Dr. Andrew Kaufman:

Thus, we can see that not only is the method that Enders created unscientific as it does not adhere to the scientific method, his conclusions were steeped in logical fallacies as well. However, these were not the only issues with Enders’ experimental method.
Enders Refuted Himself
Even though the cell culture was accepted as the “gold standard” proof for the “isolation” of a “virus,” one thing that is regularly neglected to be mentioned is that Enders himself was uncertain whether his method was even valid to begin with. In his 1954 paper, Enders questioned whether the experimental results created in a lab (in vitro) could be said to reflect what happens inside of a body (in vivo):
“The pathologic changes induced by the agents in epithelial cells in tissue culture resemble, at least superficially, those found in certain tissues during the acute stage of measles. While there is no ground for concluding that the factors in vivo are the same as those which underlie the formation of giant cells and the nuclear disturbances in vitro, the appearance of these phenomena in cultured cells is consistent with the properties that a priori might be associated with the virus of measles.“
Enders also admitted that his indirect evidence, which he used to associate with an invisible measles “virus,” was incomplete:
“Although we have thus already obtained considerable indirect evidence supporting the etiologic role of this group of agents in measles, 2 experiments essential in the establishment of this relationship remain to be carried out.”
However, the most damning revelation was the admission that during his experiments with assumed measles “virus” using the cell culture method that he invented, Enders observed the exact same cytopathogenic effects he had associated with the measles “virus” in normal control cultures without any “virus” present whatsoever:
“Monkey kidney cultures may, therefore, be applied to the study of these agents in the same manner as cultures of human kidney. In so doing, however, it must be borne in mind that cytopathic effects which superficially resemble those resulting from infection by the measles agents may possibly be induced by other viral agents present in the monkey kidney tissue (cf. last paragraph under G) or by unknown factors.“
“A second agent was obtained from an uninoculated culture of monkey kidney cells. The cytopathic changes it induced in the unstained preparations could not be distinguished with confidence from the viruses isolated from measles. But, when the cells from infected cultures were fixed and stained, their effect could be easily distinguished since the internuclear changes typical of the measles agents were not observed. Moreover, as we have already indicated, fluids from cultures infected with the agent failed to fix complement in the presence of convalescent measles serum. Obviously the possibility of encountering such agents in studies with measles should be constantly kept in mind.“
In other words, John Franklin Enders established through the use of uninoculated cultures that the cytopathogenic effect that he assumed was caused by the invisible measles “virus” occurred even when there was no assumed “virus” present within the culture. Thus, it was the cell culture process itself involving the starvation of the cell with minimal nutrients as well as the poisoning of the cell with toxic chemicals, additives, and foreign materials which eventually lead to the death of the cell resulting in the cytopathogenic effect. Therefore, it should have been concluded that the cytopathogenic effect was not the work of any fictional “virus.” This revelation should have ended virology right then and there.
However, Enders control results were instead swept under the rug and ignored. Rather than calling the time of death on virology as should have been done by anyone with an ounce of intellectual honesty, those in charge doubled down on the invalid cell culture method as the standard that every virologist must adhere to in order to confirm the presence of a “virus” within a sample. This reliance on the cell culture evidence built upon a fraudulent cytopathogenic foundation cemented the entire field of virology even further into the mad world of pseudoscience as the scientific method continued to be forsaken.
Enders Refuted by Others

Interestingly, as more researchers began to attempt to recreate Enders results, the flaws in the methodology continued to pop up. In fact, different teams of researchers over the next five years found the exact same cytopathogenic results as Enders did when they performed uninoculated controls for themselves. Dr. Cowan did an excellent overview of three of these studies in this recent video:
Let’s take a look at excerpts from the three papers Dr. Cowan discussed in order to see exactly what these researchers discovered. I’ve included highlights from each paper as well as the methods sections showing the ridiculous culturing steps each team utilized to try and “isolate” their “viruses.”
In this first study from 1955 by Rustigian et al., it is stated that the researchers discovered an unidentified “agent” within their uninoculated controls while attempting to adapt dengue “virus” to roller tube cultures of rhesus monkey kidneys. This “agent” produced the exact same cytopathogenic effect that was observed by Enders with his uninoculated culture during his measles experiments. After preparing monkey kidney cultures for a poliomyelitis study, 3 more unidentified “agents” appeared in uninoculated cultures which created the very same cytopathogenic effect. The source of these kidney cultures were healthy monkeys exhibiting no symptoms of disease, thus there was no “virus” assumed to be present within the culture. After investigation into whether or not the observed CPE could have come from the media, the researchers concluded that there was something present within the healthy monkey kidney cells that brought about the cytopathogenic effect:
Infection of Monkey Kidney Tissue Cultures with Virus-Like Agents.
“The wide current interest in tissue cultures for the study of virus-host cell relationships and applied problems of virus diseases has emphasized the need of employing cells and tissues free of microbial agents. The hazard of introducing viruses into tissue cultures with contaminated media has been stressed recently (1). Of equal, if not greater importance, is the problem of the presence of viruses in tissue cultures as a result of unrecognized infection of cells and tissues employed as primary explants. Detection of such viral infections, in certain instances, may be complicated by the fact that no readily recognized changes are manifested. On the other hand, some viruses under the same conditions may subsequently interfere markedly with cellular growth as evidenced by inhibition of acid production or
cytological changes.
In our attempts, in 1953, to adapt mouse-adapted Hawaii dengue virus(2) to roller tube cultures of rhesus monkey kidney, an unidentified agent was encountered which induced cytopathogenic changes in cultures of monkey kidney and cancer HeLa epithelial cells. The agent was filterable and could not be cultured in various non-living media. Subsequently, 3 additional agents with identical cytopathogenic characteristics were passed from uninoculated monkey kidney cultures prepared for poliomyelitis studies, to HeLa cell cultures. Enders and Peebles(3) have recently reported recovery of an agent from an uninoculated monkey kidney culture which appears to have the same cytopathogenic characteristics in monkey renal cultures as our agents have. However, other than a description of its cytopathic effect in monkey kidney cultures, no other data was given concerning their agent. In view of the wide use of monkey kidney cultures in virus studies it seems of importance to report our observations with these agents. These include their cytopathic effect in rhesus monkey renal tissue, in HeLa cell and certain other tissue cultures, the manner of their recovery and passage in tissue culture and other characteristics. In addition, evidence is presented that kidneys of apparently healthy monkeys are the source of these agents in monkey kidney tissue cultures and not the medium constituents.”
Materials and methods. Tissue culture technics. A. Roller tube cultures. Technics of Robbins, Weller and Enders (4) were essentially followed for preparation of roller tube cultures. In addition to rhesus monkey kidney, cultures were prepared frum monkey testes, human embryonic kidney, human embryonic skin and musclet and mouse infant kidney. In early studies the culture medium consisted of Hanks-Simms solution (70%) , beef embryo extract (10%) and horse serum (20%). Penicillin and streptomycin were added for final concentration of 100 units and 100 ug respectively. Later, bovine amniotic fluid was used in place of Hanks-Simms solution (5). Soybean trypsin inhibitor, in final concentration of .05 mg, was incorporated in medium for all cultures. One ml of medium was added to each culture tube containing 10 to 15 fragments of tissue. Culture fluids were changed every 3 to 4 days. At time of inoculation of agents, the medium was modified to contain 85% Hanks-Simms or bovine amniotic fluid, 10% beef embryo extract and 5% horse serum.
B. Stationary cultures of monkey kidney were prepared by trypsinization technic of Youngner(6) with bovine amniotic fluid containing 5% beef embryo extract and 10% horse serum. Following standardization of cell suspensions(7) 0.5 ml containing about 500,000 cells was added to culture tubes. When good cell growth occurred (7 to 9 days), the medium was changed to 95% bovine amniotic fluid and 5% horse serum.
C. HeLa cell strain of cancer epithelial cells, kindly supplied by Dr. J. T. Syverton of the University of Minnesota, was cultured as described by Scherer, Syverton, and Gey(8) and Syverton (9). Culture tubes were seeded with approximately 40,000 cells in 1 ml, and in 4 to 5 days a layer of growth was obtained containing 90,000 to 160,000 cells. The nutrient fluid was then replaced with maintenance solution containing 10% chicken serum(8,lO). With prolonged incubation of cultures in this fluid, degenerative changes were noted (granulation, rounding of cells). When this occurred the maintenance medium was replaced with nutrient fluid which restored normal morphology in 1 to 2 days. Accordingly, in experiments involving long incubation periods, nutrient fluid was substituted for maintenance medium and left on the cells for brief intervals.
“Recovery of agents MK1, MK3, and MK4 from uninoculated monkey kidney cultures. Shortly after encountering agent MK-D in attempts to adapt dengue virus to monkey kidney cultures, syncytial masses and vacuolatinn were again observed in an uninoculated roller tube culture 12 days after its preparation.
This culture and 16 others prepared from the same kidney were set aside for further study. By 15th day degeneration was marked in this culture and had appeared in 3 other cultures; and by 34th day in 7 of the 16 cultures. Fluids were harvested on 14th and 15th day from cultures showing degeneration, pooled and 0.2 ml inoculated into HeLa cell cultures. Lytic-like arm were present in 10 days. Fluids from these cultures were then harvested, pooled and 0.1 ml passed to fresh HeLa cultures. Similar degeneration with this second passage occurred in 6 days. Subsequent passages in HeLa cells were made with 0.1 to 0.2 ml of pooled fluids harvested at 1- to 2 day intervals for 4 to 6 days following definite signs of degeneration. The fluids were stored in a COz cabinet for varying periods between passages. This agent, designated MK1 has
now been passaged serially 101 times in HeLa cell cultures. Agents MK3 and MK4 were recovered similarly from uninoculated cultures prepared from kidneys of different rhesus monkeys. Thus, MK3 was recovered from a series of uninoculated cultures in which 8 of 72 roller tubes revealed moderate to strong degeneration 12 days after preparation and MK4 from 11 of 41 stationary cultures with degeneration 20 days after preparation with trypsinized cell suspension. These 2 agents have now been serially passed 10 and 6 times respectively in HeLa cultures. MK3 following 10 HeLa cell passages caused vacuolation and formation of syncytial masses in monkey kidney cultures. Same results were obtained with MK4 after 6 HeLa cell passages. Information has been obtained which suggests that frequency of occurrence of these agents in uninoculated cultures may be relatively high. Thus in 6 of 9 series of cultures prepared from kidneys of different monkeys one or more of 7 to 41 cultures in each series held 20 to 55 days showed characteristic degeneration (Table I); and the recovery of an agent from cultures of 3 different series was successful. One attempt to recover an agent from uninoculated cultures of a kidney series which did not show degeneration was unsuccessful after 3 blind passages in HeLa cells and subsequent passage in monkey kidney cultures. We have consistently observed in practically all cultures prepared from different monkey kidneys occasional small vacuoles apparent from the onset of cell outgrowth. In some cultures they persisted, but in others they apparently disappeared with continued incubation.”
“These findings provide evidence that the agents were derived from renal tissue of monkeys. Evidence that other tissue culture constituents are not the source of the agent is as follows: One agent was recovered at time Hanks-Simms solution was used instead of bovine amniotic fluid and the reverse was true for other 3 agents. Beef embryo extract has been routinely employed for cultivation of HeLa cells for over a year without resulting in progressive characteristic degeneration which occurred following inoculation of such cultures with the agents. Horse serum has been heated at 56°C for 30 minutes prior to its use as a medium constituent, and, as previously noted, 2 of the 4 agents so treated failed to produce cytopathic effects after 21 days. Finally 3 blind passages in HeLa cells and subsequent passage in monkey kidney cultures of fluids from one monkey kidney series failed to reveal an agent. But agents were recovered from cultures of 2 other monkey kidney series set up shortly prior to and after this series with the same lots of medium constituents.”
“Summary. During attempts to adapt dengue virus to rhesus monkey kidney cultures, an unidentified agent which causes formation of syncytial masses and vacuolation in such cultures was encountered. Subsequently, 3 additional agents with similar cytopathogenic effects were passed and maintained in HeLa cell cultures from uninoculated monkey kidney cultures. Renal tissue and not the medium constituents is the source of the agent. Bacteriological studies with one agent were negative. The same agent passed through a Selas filter. Accordingly it is considered to be virus-like in nature. Similar experiments were not done with 3 other agents but because of certain common characteristics are believed to be of the same nature.”
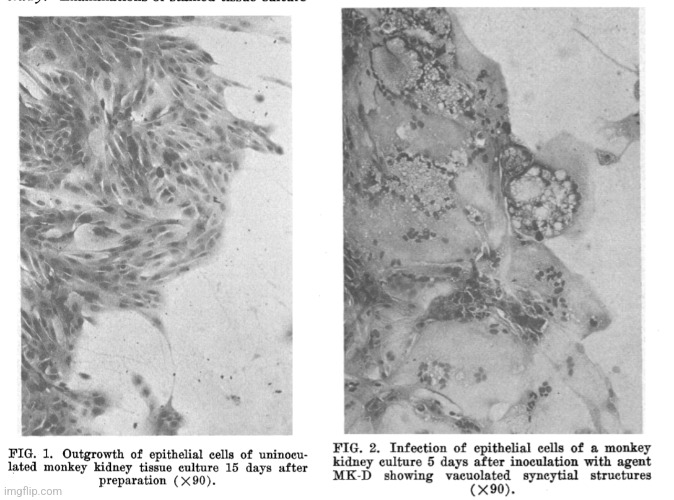
In this second study, also from 1955, Cohen et al. found the same cytopathogenic effect in their uninoculated control cultures as was observed in their inoculated samples supposedly containing a measles “virus.” Once again, this showed that the CPE was not a specific effect due to any “virus” and was therefore brought about by the experimental conditions. Cohen et al. realized that they could not use this effect to recognize the measles “virus” so they claimed that antibody results, which they admitted to presuming were due to a measles “virus,” was sufficient criteria to distinguish between the cultured samples:
Fluorescent Antibody and Complement-Fixation Tests of Agents Isolated In Tissue Culture from Measles Patients.
“Enders and Peebies(1) observed that human or monkey kidney tissue cultures inoculated with specimens from measles patients undergo, after one or more passages, characteristic nuclear and other cytologic changes. Both infectious virus and specific complement-fixation antigen appeared concurrently in the cultures and the cytopathogenic effect was neutralized by convalescent-phase measles serum. We have applied the fluorescent antibody technic of Coons and his colleagues (2,3) to similar cultures and find that the results parallel those of complement-fixation tests and thus provide immunochemical support for the data oi Enders and Peebles and in addition evidence that measles antigen(s) are present in the nuclei as well as in the cytoplasm of infected cells.
Methods. Patients were selected for study during a mild outbreak of typical rubeola. Clinical diagnosis was aided in certain cases by recognition of giant cells of the Warthin-Finkeldey type in stained films of nasal discharge obtained early in illness, as described by Tompkins and Mcaulay(4).
Collection of specimens and preparation of tissue cultures. The procedures outlined by Enders and Peebles (1) were followed. Blood, throat swabs or washings, or nasal discharge were obtained before or within a day after the appearance of the rash. When possible, specimens were inoculated into tissue cultures within a few hours after their collection. Tissue cultures consisted of trypsinized monkey kidney cells grown in sheets (5). Tubes were inoculated with 0.2 or 0.3 ml of the prepared specimens, fed with nutrient fluid to bring the volume to 1.0 ml, and incubated as stationary slants at 36.5°C. Nutrient fluid consisted of beef amniotic fluid containing 5% inactivated horse serum and 5% beef embryo extract. The first passage contained 100 units of penicillin, 100 ug of streptomycin, and 50 ug of nystatin (fungicidin) (6): others, 50 units and 50 and 10 ug, respectively. Transfers to fresh tubes were carried out at 14-day intervals with 0.2-ml aniounts of pooled fluids taken 5, 9, and 14 days after inoculation. Controls consisted of uninoculated cultures and of cultures passaged serially with fluids from uninoculated tubes.
Results. Transmissible agents, presumably viruses, were isolated from both blood and a throat swab of one patient, and from nasal discharge and a throat swab, respectively, of 2 others whose blood was not obtained. The throat swab specimens had been stored in a dry ice chest for 7 days. No agents were detected in specimens from 5 additional cases. Inocula from 3 of the latter had been stored at 4°-6°C for 2 days before culture; tests of inocula from the other 2 were discontinued after 3 apparently negative serial passages. These circumstances may have contributed to our failure to isolate agents.
Enders and Peebles(1) and Rustigian et al. (10) encountered latent virus-like agents that induce marked vacuolization and syncytial masses in monkey kidney tissue cultures. The cellular degeneration characteristic of these “monkey-kidney agents” frequently appeared in our cultures, both in those inoculated with specimens from measles patients and in controls; hence cytologic criteria for recognition of measles agents were difficult to apply. In some tissue culture series the “monkey-kidney agents” destroyed the cell sheets in 10 to 14 days. We therefore relied on tests for the presence of measles antigen to identify the 3 agents cultivated from measles inocula.”
“Discussion. Demonstration that tissue cultures inoculated with specimens from measles patients produce antigens that react specifically with convalescent-phase measles sera substantiates the findings of Enders and Peebles( 1). We did not repeat their filtration experiments, but were unable to recover fungi, nodes from rabbits immunized with diphtheria or bacteria including pleuropneumonia-like organisms and leptospira from infected tissue cultures and presume, therefore, that the antigens that produced specific reactions in fluorescent antibody and complement-fixation tests were derived from measles virus. On the whole, antigen in infected tissue cultures was detected earlier by the fluorescent antibody method than by complement-fixation tests. Preliminary data suggest, however, that the latter are of value in differential diagnosis of exanthemata and encephalitides of unknown origin.”
This final study which Dr. Cowan highlighted is from Von Magnus et al. in 1959. In lockstep with the previous researchers, they also came across the same exact cytopathogenic effect in uninoculated cultures from healthy monkeys. The researchers claimed that the changes were due to what they referred to as “foamy agents” and stated that these “agents” are regularly found in healthy monkey kidney cells. The CPE generated was indistinguishable from that which was associated with the measles “virus.” Thus, like Cohen et al. before them, Von Magnus et al. decided that this effect was of limited use in attempting to identify “viruses” and instead relied upon unreliable antibody tests to determine whether the measles “virus” was present in a sample. Also of interest is that these researchers failed in numerous attempts to “isolate” the measles “virus” as they were only “successful” in 5 of 13 cases. All attempts to isolate a “virus” failed 24 hours after onset of a rash. Regardless of their failures, Von Magnus et al. decided that their indirect evidence agreed with the assumption that the antibody results identified the measles “virus:”
Studies on Measles Virus in Monkey Kidney Tissue Cultures. 1. Isolation of Virus from 5 Patients with Measles.
“As described by Enders & Peebles (6), and later by Rustigian et al. (13) and by Cohen el al. (3) cytopathic changes similar to those caused by measles virus may be observed also in uninoculated cultures of monkey kidney tissue (Figs. 4-5). These changes are probably caused by virus-like agents, so called “foamy agents”, which seem to be frequently present in kidney cells from apparently healthy monkeys. Specific measles antigen is, however, produced only in cultures infected with measles virus. Iu the present study the ability of tissue culture passage material to fix complement in the presence of convalescent phase measles serum was therefore used as a criterion for the presence of measles virus.”
MATERIALS AND METHODS
Tissue Cultures: Cell of trypsinized kidneys from rhesus monkeys, cynomolgous monkeys or baboons were u sed routinely. The technique of Younger (15) for the preparation or these tissue cultures was used with some modifications as previously described (10). During the outgrowth of the cells, the growth medium consisted of lactalbumln hydrolysate 0.5 cent (11) in Hank’s solution (5) with 2 per cent horse serum.
Before seeding with the virus the medium was changed. Each tube received 1.8 ml of either synthetic medium 199 (12) or bovine amniotic fluid (5) containing phenol red as an indicator (final dilution 0.02 per cent). All media contained penicillin (100 U/ml) and streptomycin (0.1 mg/ml).
Collection of specimens: isolation of the virus was attempted from throat washings and blood.
Throat washings: Measles patients were askcd to gargle either with 15 ml of a mixture of one part ox-heart infusion broth and 2 parts of buffered salt solution or, in later experiments, with 15 ml of distilled water containing l percent Bacto tryptose “Difco”. Penicillin, 100 U. per ml and streptomycin 0.1 mg per m l were added to the fluid. From the throats of very young children specimens were obtained by cotton swabs which were subsequently immersed into 2 ml of one of the two fluids just described. In all instances, the specimens were immediately froten in C02-ice and then stored in an electrical deep-freeze (-60° C). Before inoculation into tissue culture the materials were thawed rapidly at 37°C in running tap water.
Blood: ln earlier experiments heparin was added to the blood (2 ml of an O.O5 per cent of heparin per 10 ml of blood). In later experiments the blood was allowed to coagulate. Red cells which had not become attached to the clot were resuspended In the serum, and this mixture was used as inoculum for tissue cultures. The blood samples were stored at +4° C until the time of inoculation.
Inoculation of tissue cultures: Tissue cultures were inoculated with 0.5 rnl of throat washing material or with 0.25 ml of blood. The final amount of fluid in the tubes was about 2 ml. The cultures were kept staUonary or placed in a rotating drum (1 rotation per minute).
Subcultures from the first passage were carried out between the 6th and the 16th day, usually on the 8th day after inoculation. The nutrient medium was as a rule not exchanged in the course of a passage. Subcultures from the later passages were made between the 6th and the 12th day of incubation. The passage material consisted of a suspension of cells and cell debris in culture medium. This mixture was obtained by loosening the cells still adhering to the glass through scraping with a pipette. The amount inoculated varied, but it was usually 0.2 mi, so that the final volume of fluid in the inoculated tubes was about 2 ml. Serial passages of material from control tubes were carried out in the same way.
DISCUSSION
“The present work confirms the findings of Endel’s & Peebles (G) and Cohen et al. that throat washings and blood frotn patients in the early stage of measles contain a virus which is able to produce characteristic cytopathic changes in cultures of human and simian kidney cells. In the inoculated cultures the same syncylical formations were observed as those described by Enders & Peebles (6). In addition it was found that in the tissue culture system employed in this other specific lesions developed on continued incubation. This second step of the degeneration resulted in the accumulation of cell debris with circular gritty-looking formations ,vhich had either a quite smooth or a wrinkled margin. These cytopathic changes appear to be as specific for measles virus as are the synctia.
However, monkey kidney viruses or “foamy agents,” may give rise to cellular degenerations which microscopically are indistinguishable from those caused by measles virus. For this reason the cytologic manifestations are of limited value in the study of measles and additional criteria ate required to establish the identity of the cultivated agents. This can be achieved by the demonstration of intranuclear inclusion bodies in the cell cultures (6) or by tests for the presence of measles anligen using either the fluorescent antibody technique (3) or the complement fixation reaction (3, 6). Because of its simplicity this last mentioned method has been employed in the present study. Observations on the development of measles antigen in the various cultures will be described in detail in a forthcoming publication (1).
In the study presented in this paper measles virus was isolated from five out of nine throat washings collected withln 24 hours after the outbreak of exanthem. The reason why the isolation failed in one patient (No. 12) may be that he had a vomiting immediately before gargling. In the remaining three cases (Nos. 2, 7 and 9) no obvious explanation for the negative results can be offered. Possibly it was because these isolations were all attempted with throat swabs in young children who were very excited and the sampling thus difficult to perform.
Virus was recovered from the blood in only one out of eight attempts made within 24 hours after the onset of the rash. This isolation rate is low compared with that obtained by Enders & Peebles (6) who recovered virus from the blood of four out of five patients. These authors used heparinized blood from which it may be easier to recover virus than from serum containing resuspended blood cells. In our laboratory the only isolation from the blood was made from one of the three samples to which heparin had been added. Also, Enders and Peebles used a larger inoculum (0.5 ml to 2 ml) than employed in this study (0.25 ml).
The possibility was considered that the failure to recover virus from 8 of the 13 patients examined might be due to an insusceptibility to measles virus of the particular cells employed. This, however, was apparently not the case, for when tubes prepared simultaneously with those employed in the unsuccessful isolation experiments were inoculated with measles infected fluid typical cytopathic changes developed together with the appearance of complement-fixing antigen in the nutrient media.
The observations presented in this paper agree well with the assumption that the isolated agents are the cause of measles.”
SUMMARY
(1) Virus agents have been isolated in trypsinized monkey kidney tissue cultures from throat washings and blood from 5 out of 13 patients examined during the acute phase of measles. In all instances, virus was isolated from throat washings or throat swahs while one strain was recovered from the blood. All attempts to isolate virus later than 24 hours after onset of the rash failed.
2) The cytopathic manifestations observed in measles virus infected tissue cultures as well as in uninoculated tubes are described. Complement fixation tests for the presence of measles antigen have been used as criterion for the presence of this virus in the infected cultures.
(3) Intranasal and oral administration of material from late passages of one of the isolated agents to two Rhesus resulted in a pronounced measles-like rash in one of the animals, and both monkeys developed antibodies against the inoculated strain.
(4) In serological studies of acute and convalescent-phase sera from measles patients using measles virus infected cultures of monkey kidney tissue as antigen in complement fixation tests a clearcut rise in antibodies was observed in all cases.

What we can take away from these three studies in combination with Enders’ own uninoculated controls is that no “virus” was ever necessary in order for the cytopathogenic effect to occur. This effect was witnessed in cultures of kidney cells from healthy monkeys. It was stated that these findings were not rare as witnessing this cytopathogenic effect in uninoculated cultures from healthy monkeys was actually rather frequent. Thus, it is clear to see that it is the cell culture method itself, which consists of stressing and starving the cells which are outside of their natural environment and dousing them in kidney toxic antibiotics as well as mixing in foreign animal materials and other chemical additives that leads to the death of the cell. As this cytopathogenic effect was not specific to a “virus” and occurred in cultures without any “viruses” present, it can not be used to claim the presence of a “virus” as both Cohen et al. and Von Magnus et al. stated. Thus, there is no value to any results gained from this pseudoscientific experimentation.

While the above studies should be enough to wash away any doubt left about the fraudulent use of a cytopathogenic effect as confirmation that a “virus” is present in the sample, I am throwing a 1956 publication by Hull et al. into the mix as well which provides even more damning evidence supporting the discontinuation of this pseudoscientific practice. In this paper, the researchers stated that the use of the cell culture method led to the discovery of many unknown cytopathogenic agents that can only be detected through the use of this practice. The researchers used slight variations in the CPE observed during the culture for poliomyelitis vaccines to claim that there were at least 10 other invisible “virus-like agents” present at various times within the cultures. While Hull et al. believed that these agents most likely originated within the monkey kidney cells, they said there was a potential that these “agents” could represent contaminants from human sources, horse serum, nutrient medium or other solutions used in the preparation of the cultures. Regardless, they decided that these invisible agents, which were “recovered” from normal or uninoculated control cultures from healthy monkeys, were “simian viruses.” The researchers did so despite the fact that they were unable to make any animals sick through various routes of injection beyond two exceptions where two of the “viruses” caused death upon injection in the brain. However, upon injection of these two “viruses” into the muscles, no illness occurred. In other words, Hull et al. observed cytopathogenic effects that they subjectively determined were not caused by the polio “virus” in both inoculated and uninoculated cultures and decided that there must be different “viruses” present within the culture based on the pattern of cell death observed. These mixtures assumed to contain invisible “viruses” were consistently demonstrated not to be pathogenic, hence the unobservable entities from uninoculated cultures with the same pattern of CPE from healthy monkeys did not meet the definition of a “virus:”
Viral Agents Recovered from Tissue Cultures of Monkey Kidney Cells
“Increased use of the technique of cell cultivation for isolation, maintenance and study of viruses has resulted in the discovery of many hitherto unknown cytopathogenic agents. In screening human stool samples or rectal swabs for poliomyelitis viruses, Melnick (1) isolated several “orphan viruses” which do not type with poliovirus antiserums, and from the same source Sabin (2) obtained 5 viruses referred to as He1; He2, etc. The new RI series of viruses was isolated in tissue culture by Hilleman and Werner (3) from throat washings of patients with respiratory disease. Rowe, Huebner et al. (4) isolated a similar group of viruses, the APC group, which first appeared in tissue cultures of human adenoid and tonsillar tissue and later were recovered from eye washings of patients with a conjunctivitis. Recently Rustigian et al. (5) reported recovery of two cytopathogenic agents from cultures of monkey kidney cells. Thus, a number of viral agents are present in tissues and excreta of man and lower animals which defy detection by methods other than tissue culture.
During production and testing of poliomyelitis vaccine, hundreds of thousands of monkey kidney cultures prepared from thousands of monkeys were observed in our laboratories and in others participating in this program. Numerous filterable, transferable cytopathogenic agents other than poliovirus were also encountered. Although these agents were probably introduced into the cultures with the monkey kidney cells, there remains the remote possibility that some represent contaminants from human sources, horse serum, nutrient medium or other solutions used in the preparation of cultures. This report describes 8 immunologically distinct agents isolated and studied in our laboratories, and makes reference to 2 from other sources.”
“The agents isolated will be referred to as “Simian viruses” (S.V.) until such time as a definite association with some other host or identification can be established. These agents will be referred to as S.V.1, S.V.2, etc. Of the original group of agents which included S.V., through S.V.15, eight (S.V. 1, 2 4, 5. 6, 11, 12 and 15) will be described in this paper. The missing numbers were held by agents that were identified or reclassified after antiserum studies, or which have not yet been adequately studied for inclusion at this time. The latter category includes S.V.13 which is
the “lacy or foaming” agent frequently seen in cultures of monkey kidney cells.
RESULTS
The following is a brief account of the original isolation and description of the 8 agents referred to above. S.V.1 was recovered from roller-tube cultures of trypsinized monkey kidney cells planted on February 8, 1954. This series included 1,700 tubes prepared from a pool of kidneys from 4 apparently healthy cynomolgus monkeys. After 6 to 8 days of incubation the cultures appeared satisfactory and were used for titrations of poliovirus. After 6 more days of incubation a type of cell destruction was seen in some cultures beyond the end point of the poliovirus that was atypical of the damage produced by the latter. This atypical damage was observed in 17 percent of the cultures. This eytopathogenic effect (C.P.E.) was characterized by a reduction in size and distinct rounding of the cells.”
“Monkeys were inoculated intracerebrally, intramuscularly, and in some instances by other routes, with all the viruses. Serum from each monkey inoculated was first assayed for antibodies against the specific virus to be injected. Only negative animals were used. With the exception of S.V.12 and S.V.15 no evidence of clinical illness or of gross or histopathological lesions was seen. Monkeys inoculated intracerebrally with high titer undiluted S.V.12 and S.V.15 viruses succumbed within 4 to 6 days after inoculation. The specific type of virus inoculated was readily recovered from brain and cord tissues of these animals. These same two viruses, however, when inoculated intramuscularly into monkeys failed to produce clinical disease or gross or histopathological lesions. The histopathological lesions observed in the intracerebrally inoculated monkeys revealed necrosis and complete destruction of the choroid plexus plus a generalized aseptic type of meningitis. No other brain pathology or lesions resembling those of poliomyelitis were seen. The various organs studied exhibited no recognized lesions and it appeared that all involvement was limited to the central nervous system in the region of the virus inoculation. The lesions produced by S.V.12 and S.V.I5 were indistinguishable.
Although no recognizable disease was produced in the monkeys inoculated with these viruses, with the above noted exceptions, some evidence was obtained in our laboratories, and in others, that S.V.2 and S.V6, may have some etiological significance in enteric infections.”
“None of these monkey viruses infected embryonated eggs as determined by death of the embryo. However, one agent from the A.M.S.G.S. identified as S.V., had been recovered from embryonated eggs inoculated with monkey kidney tissue-culture fluids. Groups of adult and suckling mice were inoculated intracerebrally and intraperitoneally with each virus. However, no evidence of disease nor death of the mice was observed. Rats and rabbits which were inoculated with live virus for the production of antiserum also did not succumb to infection with any of these agents.”
DISCUSSION
The isolations and characteristics of 8 apparently new viruses have been described. Although definite proof has not been presented, these agents probably have been simian in origin. What diseases, if any, they might be responsible for in monkeys also has not been established. S.V.2 and S.V. 6, may be etiological agents in intestinal infections of monkeys inasmuch as both have been isolated from stool samples of animals suffering from diarrhea. None have produced experimental infections when inoculated intramuscularly although both S.V.12 and S.V.15 caused a prostrating type of paralysis and death in monkeys following intracerebral inoculation. Attempts which are being made to infect monkeys by other routes of inoculation are not yet completed.”
The greatest significance of these viruses has been their appearance in tissue cultures used to produce and to test poliomyelitis vaccine. This has been especially troublesome during tissue-culture safety testing of the vaccine as the occurrence of these viruses has in many instances invalidated tests for poliovirus, making retesting necessary and thereby delaying the release of vaccine. There has also been the problem of proving in some cases that the contaminating ” simian virus” isolated during a vaccine safety test was not in the vaccine sample under test. In general, however, this has not been a problem since many of these agents have been recovered from normal or uninoculated control cultures.”
SUMMARY
“Eight apparently new submicroscopic, filterable, cytopathogenic agents recovered in tissue cultures of rhesus and cynomolgus monkey kidney tissue have been described. No definite associations have been made between any of these agents and naturally occurring diseases of monkeys or other animals. The significance of these viruses in the poliomyelitis vaccine program and the problems created by them during vaccine safety testing in tissue culture have been discussed.”
These studies in and of themselves should be enough to show that, when controls are done, the cytopathogenic effect observed and attributed to the presence of an invisible “virus” is shown not to be specific to any “virus.” In fact, no “viruses” are necessary to create this effect whatsoever. Luckily, we are fortunate enough that Dr. Stefan Lanka did not just rest on the work of others to show this and took it upon himself to verify these findings by doing his own set of control experiments, thus putting the proverbial final nail in the coffin of this pseudoscientific practice. As Dr. Cowan did an absolutely excellent job breaking down Dr. Lanka’s control experiments, I am providing excerpts from his book Breaking the Spell:

“Here is the essence of Lanka’s experiment, done by an independent professional laboratory that specializes in cell culturing. As seen in this series of photographs, each of the four vertical columns is a separate experiment. The top photo in each column was taken on day one, and the bottom photo was taken on day five.
In vertical column one, normal cells were cultured with normal nutrient medium and only a small amount of antibiotics. As you can see, on neither day one nor day five was any CPE found; the cells continued their normal, healthy growth.
In vertical column two, normal cells were again grown on normal nutrient medium and a small amount of antibiotics, but this time, 10% fetal calf serum was added to enrich the medium. Still, the cells in the culture grew normally, both on day one and day five.
The third vertical column shows what happened when Dr. Lanka’s group used the same procedures that have been used in every modern isolation experiment of every pathogenic virus that I have seen. This included changing the nutrient medium to “minimal nutrient medium”—meaning lowering the percentage of fetal calf serum from the usual 10% to 1%, which lowers the nutrients available for the cells to grow, thereby stressing them—and tripling the antibiotic concentration. As you can see, on day five of the experiment, the characteristic CPE occurred, “proving” the existence and pathogenicity of the virus—except, at no point was a pathogenic virus added to the culture. This outcome can only mean that the CPE was a result of the way the culture experiment was done and not from any virus.
The fourth and final vertical column is the same as vertical column three, except that to this culture, a solution of pure RNA from yeast was added. This produced the same result as column three, again proving that it is the culture technique—and not a virus—that is causing the CPE.”
Invalid Experiment = Invalid Controls

It should hopefully be clear by now why virology has a control problem. The cell culture method itself is not a valid experimental set-up as it was never designed through the adherence to the scientific method. The experiment creates the effect (CPE) and then assumes a cause (“virus”) without ever verifying that the assumed cause exists in the first place. Even if we were to give them leeway with the use of the cell culture as a valid experiment to prove the presence of a “virus,” the cytopathogenic effect is known to be caused by many other factors unrelated to any “virus” thus making the explanation of a fictitious “virus” as the culprit entirely unnecessary. The blaming of invisible “viruses” for observed CPE in place of other explanations was pointed out in a technical bulletin for cell culturing:
Understanding and Managing Cell Culture Contamination
“Since cytopathic viruses usually destroy the cultures they infect, they tend to be self-limiting. Thus, when cultures self-destruct for no apparent reason and no evidence of common biological contaminants can be found, cryptic viruses are often blamed. (See Figures 3a and 3b.) They are perfect culprits, unseen and undetectable; guilty without direct evidence. This is unfortunate, since the real cause of this culture destruction may be something else, possibly mycoplasma or a chemical contaminant, and as a result will go undetected to become a more serious problem.”

It is known that the CPE can be caused by factors other than “viruses” such as:
- Bacteria
- Amoeba
- Parasites
- Antibiotics
- Antifungals
- Chemical contaminants
- Age and cell deterioration
- Environmental stress
And lest I forget, the also fictional “virus-like” entities (that are admitted to be impossible to separate from “viruses”) known as exosomes are also said to create cytopathogenic effects.

To reiterate, the cytopathogenic effect is not a valid dependent variable as it is not a naturally observed phenomena, and it can be explained by various factors other than an invisible “virus.” The unpurified fluids used during the culture is not a valid independent variable as the “virus” assumed to be within has never been shown to exist in a purified and isolated state before the experiment takes place. Thus, performing the cell culture as evidence for a “virus” is entirely unscientific as the scientific method can not be followed. However, for arguments sake, let’s allow virologists to have the cell culture and the cytopathogenic effect as a valid experiment. This would mean that they would need to have valid controls performed alongside the cell cultures every time. One would think that this would be commonly seen in virology papers as we saw in those from the 1950s. Yet, more often than not, either no mention of the controls are ever found within the studies provided as evidence for the existence of “viruses” or what was done to the control culture is ill-defined. Perhaps this is due to the disastrous conclusions made by the authors of the 1950’s studies, which irreparably damaged the claim of a “viral” cause of the CPE observed? In any case, if virologists do perform a control, they usually do what they refer to as mock infections:
mock-infected
“A control used in infection experiments. Two specimens are used one that is infected with the virus/vector of interest and the other is treated the same way except without the virus. Sometimes a non-virulent strain is used in the mock-infected specimen.”
https://www.genscript.com/biology-glossary/10558/mock-infected
What this means is that the virologists are supposed to use the same cell with the same additives (antibiotics, antifungals, minimal nutrient media, fetal bovine serum, etc.) but without the “virus” added in. Let’s take a look at a few examples in order to see if this is what they do. In this first instance, the authors of a CDC study state that the mock-infected culture was treated with the medium only, so this appears to match up to the above definition. However, it is not stated whether the medium was exactly the same as that of the experimental culture. We must assume this to be the case, which is a problem as will be shown later.
SARS–associated Coronavirus Replication in Cell Lines

This second study is one of the pivotal early “SARS-COV-2” studies by Zhu et al. We are only told that the control is mock-infected and thus we must once again assume that the researchers used exactly the same medium/ingredients as no other details were provided.
A Novel Coronavirus from Patients with Pneumonia in China

This third example is from an Australian study in April 2020 which is infamous for adding trypsin, a protein digester, to the culture in order to create the “spike” corona in EM images. The researchers give us what they call uninfected control cells. Again, we must assume that the uninfected cell was treated exactly the same as the infected cell as no details were provided.
Isolation and rapid sharing of the 2019 novel coronavirus (SARS-CoV-2) from the first patient diagnosed with COVID-19 in Australia

In this final study from Zhou et al., which is one of the pillars of the “SARS-COV-2” fraud, we are told that a mock “virus” was used. What is a mock “virus?” Zhou et al. do not say.
A pneumonia outbreak associated with a new coronavirus of probable bat origin
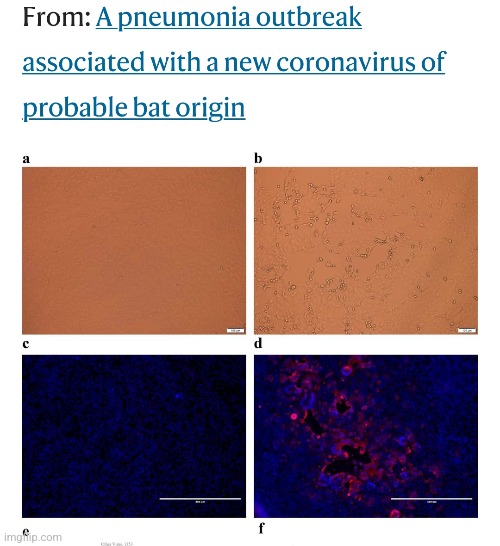
Or they did not say so in public. However, in a private correspondence through e-mail, some further details were shed on this situation. From Dr. Mark Bailey’s amazing essay “A Farewell to Virology,” we find out that in the experimental culture, the antibiotics were doubled during the culture experiments to achieve a cytopathogenic effect in 1 out of 24 cultures. Not only is this a stunning failure rate to culture a “virus,” the addition of more antibiotics to the experimental culture completely invalidates the results as the control was not treated the same.

As can be seen in these examples, what was done to the mock-infected controls is not well-defined and must be taken as an assumption that the two cultures were treated the same minus the assumed “virus.” However, the Zhou et al. admission is exactly why we can not assume that the cultures are treated equally as this was obviously not the case. The addition of more antibiotics to the experimental culture was never mentioned anywhere within the paper. Zhou et al. committed scientific fraud. How many other “virus” studies would be shown to have done the same if the researchers were as honest as Xing-Lou Yang was in an e-mail exchange? The details of what was done to the mock infected controls must be provided with every paper yet this is rarely, if ever, the case.
However, there is an even bigger underlying problem for virology than a lack of ill-defined mock-infection controls even if all of the same additives are used. Remember, a control is supposed to eliminate only the one variable under study, i.e. the assumed “viral” particles. As the fluids that are utilized for the “inoculated” culture are admitted to not contain only purified and isolated “virus” particles but a whole gamut of potential substances such as host materials, bacteria, fungi, microvesicular bodies, etc., mock-infections where no human fluids are added to the culture are not proper controls. A proper control would be to use a sample from a healthy human which is treated in the exact same manner as the fluids with the assumed “virus.” This includes adding the healthy human fluids to “viral” transport media which contains added chemicals, nutrients, fetal bovine serum, antibiotics/antifungals, etc. as this step is done immediately to the “viral” sample upon collection. Leaving fluids from healthy subjects out of the control invalidates the mock-infection as there are numerous confounding variables present within the experimental culture that are missing from the mock-infected culture. Thus, while a mock-infected control can be said to be a control, it is not the proper control for the pseudoscientific cell culture experiments.

In Summary:
- A study with control(s) is designed to ensure that the effects are due to the independent variables in the experiment
- The use of controls allows to study one variable or factor at a time
- It is important that both the control and other (experimental) group(s) are exposed to the same conditions apart from the one variable under study
- Relating to virology, that one variable would be the assumed “viral” particles and thus it is imperative that since virologists use an unpurified sample from sick humans, they would need to use an unpurified sample from a healthy human as a control as well
- Throughout the early 20th century, very different interpretations of the nature of “viruses” arose, which were put forward against each other
- No experimental evidence for this or that concept, which all researchers should have accepted, could be presented by any side
- Findings often reported by certain “virus” researchers at the time were not confirmed by other researchers as a result of their own experiments, or the observations could not be reproduced by all scientists working with the “virus”
- Often, findings to the contrary were reported, or the findings that had been examined were considered artefacts
- Every version of the interpretation of the “virus” phenomenon remained open to attack
- Facts presented to the expert public could often be reinterpreted into fictions by opponents, who brought into play the dependence of the findings on the conditions of observation, the local situation of the experiments, the research-related nature of the attributions of characteristics, etc. as sources of error
- Findings that were used to empirically confirm a suspected connection were often soon joined by negative findings reported by other researchers
- At no time did one party offer compelling reasons that would have led the other party to finally abandon artifact accusations
- In 1954, the tissue culture practice, which had been shown to produce the same results in healthy tissues, fell out of favor for John Franklin Enders new cell culture method
- The cell culture relied on the creation of the cytopathogenic effect (CPE), patterns of cell death, as being specific to “viruses” in order to identify that they were present within the culture
- Even though this method was quickly accepted as the new gold standard for “virus isolation,” Enders himself questioned his results as he observed indistinguishable cell death in the healthy control cultures as seen in those with the measles “virus”
- According to Enders, a second agent was obtained from an uninoculated culture of monkey kidney cells
- The cytopathic changes it induced in the unstained preparations could not be distinguished with confidence from the “viruses” isolated from measles
- He felt it was obvious that the possibility of encountering such agents in studies with measles should be constantly kept in mind
- Other researchers over the next 5 years came to the same conclusions as they also observed the same cytopathogenic effect in their own healthy control cultures

- The hazard of introducing “viruses” into tissue cultures with contaminated media has been stressed recently
- Detection of such “viral” infections, in certain instances, may be complicated by the fact that no readily recognized changes are manifested (a.k.a. the CPE-less “viruses” escape clause)
- In 1953, the researchers attempted to adapt mouse-adapted Hawaii dengue “virus” to roller tube cultures of rhesus monkey kidney and an unidentified agent was encountered which induced cytopathogenic changes in cultures of monkey
kidney and cancer HeLa epithelial cells - Three additional agents with identical cytopathogenic characteristics were passed from uninoculated monkey kidney cultures prepared for poliomyelitis studies, to HeLa cell cultures
- Enders and Peebles reported recovery of an agent from an uninoculated monkey kidney culture which had the
same cytopathogenic characteristics in monkey renal cultures as their agents - Evidence was presented that kidneys of apparently healthy monkeys are the source of these agents in monkey kidney tissue cultures and not the medium constituents
- Shortly after encountering agent MK-D in attempts to adapt dengue “virus” to monkey kidney cultures, syncytial masses and vacuolatinn were again observed in an uninoculated roller tube culture 12 days after its preparation
- Agents MK3 and MK4 were recovered similarly from uninoculated cultures prepared from kidneys of different rhesus monkeys
- Information was obtained which suggested that the frequency of occurrence of these agents in uninoculated cultures may be relatively high
- The researchers consistently observed in practically all cultures prepared from different monkey kidneys occasional small vacuoles apparent from the onset of cell outgrowth
- The researchers stated that these findings provided evidence that the agents were derived from renal tissue of monkeys and that it was the renal tissue and not the medium constituents which was the source of the agents

- Enders and Peebies observed that human or monkey kidney tissue cultures inoculated with specimens from measles patients undergo, after one or more passages, characteristic nuclear and other cytologic changes
- In cell culture experiments for ruebella by Cohen et al., controls consisted of uninoculated cultures and of cultures passaged serially with fluids from uninoculated tubes
- Transmissible agents, presumably “viruses,” were said to be isolated from both blood and a throat swab of one patient, and from nasal discharge and a throat swab, respectively, of 2 others whose blood was not obtained
- No agents were detected in specimens from 5 additional cases
- Inocula from 3 of the latter had been stored at 4°-6°C for 2 days before culture; tests of inocula from the other 2 were discontinued after 3 apparently negative serial passages and Cohen felt that these circumstances may have contributed to their failure to isolate agents
- Cohen reiterated that Enders and Peebles and Rustigian et al. encountered latent “virus-like” agents that induce marked vacuolization and syncytial masses in monkey kidney tissue cultures
- Keep in mind that they did not observe any “virus-like” agents, they just assumed that they must be present as the same CPE was observed in controls said to contain no “virus”
- The cellular degeneration characteristic of these “monkey-kidney agents” frequently appeared in our cultures, both in those inoculated with specimens from measles patients and in controls; hence cytologic criteria for recognition of measles agents were difficult to apply
- In other words, the CPE was identical as they could not tell any difference between the CPE observed in the uninoculated controls and the inoculated samples and could not use CPE as criteria to recognize the measles “virus”
- Cohen et al. did not repeat Enders filtration experiments, but were unable to recover fungi, nodes from rabbits immunized with diphtheria or bacteria including pleuropneumonia-like organisms and leptospira from infected tissue cultures so they presumed that the antigens that produced specific reactions in fluorescent antibody and complement-fixation tests were derived from measles “virus”
- In other words, as they could not use CPE to determine measles cases, they used non-specific antibody results which they presumed were specific to a measles “virus”

- As described by Enders & Peebles, and later by Rustigian et al. and by Cohen el al. cytopathic changes similar to those caused by measles “virus” may be observed also in uninoculated cultures of monkey kidney tissue
- These changes are probably caused by virus-like agents, so called “foamy agents”, which seem to be frequently present in kidney cells from apparently healthy monkeys
- This second step of the degeneration resulted in the accumulation of cell debris with circular gritty-looking formations , which had either a quite smooth or a wrinkled margin (cell-debris with a circular formation sure sounds a lot like a “virus” 🤔)
- Monkey kidney “viruses” or “foamy agents,” may give rise to cellular degenerations which microscopically are indistinguishable
from those caused by measles “virus” - They decided for this reason the cytologic manifestations are of limited value in the study of measles and additional criteria are required to establish the identity of the cultivated agents
- In the study presented in this paper measles “virus” was isolated from five out of nine throat washings collected within 24 hours after the outbreak of exanthem
- The reason why the isolation failed in one patient was thought to be because he had vomited immediately before gargling
- In the remaining three cases, no obvious explanation for the negative results can be offered but they thought it may possibly be because these isolations were all attempted with throat swabs in young children who were very excited and the sampling thus difficult to perform
- “Virus” was recovered from the blood in only one out of eight attempts made within 24 hours after the onset of the rash which was low compared with that obtained by Enders & Peebles who recovered “virus” from the blood of four out of five patients
- The possibility was considered that the failure to recover “virus” from 8 of the 13 patients examined might be due to an insusceptibility to measles “virus” of the particular cells employed but this theory was abandoned
- They stated that the observations presented in this paper agreed well with the assumption that the isolated agents were the cause of measles
- All attempts to isolate “virus” later than 24 hours after onset of the rash failed
- The cytopathic manifestations observed in measles “virus” infected tissue cultures as well as in uninoculated tubes were described
- Intranasal and oral administration of material from late passages of one of the isolated agents to two Rhesus resulted in a pronounced measles-like rash in one of the animals, and both monkeys developed antibodies against the inoculated strain

- Hull et al. stated that the increased use of the technique of cell cultivation for isolation, maintenance and study of “viruses” has resulted in the discovery of many hitherto unknown cytopathogenic agents
- Various researchers had found a number of “viral agents” which are present in tissues and excreta of man and lower animals which defy detection by methods other than tissue culture
- During production and testing of poliomyelitis vaccine, hundreds of thousands of monkey kidney cultures prepared from thousands of monkeys were observed in our laboratories and in others participating in this program and numerous filterable, transferable cytopathogenic agents other than “poliovirus” were also encountered
- Hull stated that although these agents were probably introduced into the cultures with the monkey kidney cells, there remained the remote possibility that some represent contaminants from human sources, horse serum, nutrient medium or other solutions used in the preparation of cultures
- The agents isolated were referred to as “Simian viruses” (S.V.) until
such time that a definite association with some other host or identification could be established - One of these included S.V.13 which is the “lacy or foaming” agent frequently seen in cultures of monkey kidney cells discussed in the studies above
- For S.V. 1, 1,700 tubes prepared from a pool of kidneys from 4 apparently healthy cynomolgus monkeys were used
- After 6 to 8 days of incubation the cultures appeared satisfactory and were used for titrations of “poliovirus” and after 6 more days of incubation a type of cell destruction was seen in some cultures beyond the end point of the “poliovirus” that was atypical of the damage produced by the latter
- In other words, a longer incubation period resulted in further cytopathogenic effects which were (subjectively) considered not due to polio so they determined it must have been caused by another “agent”
- This atypical damage was observed in 17 percent of the cultures
- Monkeys were inoculated intracerebrally, intramuscularly, and in some instances by other routes, with all the “simian viruses” and with the exception of S.V.12 and S.V.15, no evidence of clinical illness or of gross or histopathological lesions was seen
- Monkeys inoculated intracerebrally with high titer undiluted S.V.12 and S.V.15 “viruses” succumbed within 4 to 6 days after inoculation
- These same two “viruses,” however, when inoculated intramuscularly into monkeys failed to produce clinical disease or gross or histopathological lesions
- It can be seen the assumed S.V.’s showed no signs of pathogenicity at any point, except for S.V. 12 & 15 which killed monkeys upon injections in the brain but did nothing upon injections in the muscles, thus they are not “viruses”
- No recognizable disease was
produced in the monkeys inoculated with these “viruses” - None of these monkey “viruses” infected embryonated eggs as determined by death of the embryo
- Groups of adult and suckling mice were inoculated intracerebrally and intraperitoneally with each “virus” and yet no evidence of disease nor death of the mice was observed
- Rats and rabbits which were inoculated with live “virus” for the production of antiserum also did not succumb to infection with any of these agents
- The “isolations” and characteristics of 8 apparently new “viruses” have been described yet no definite proof had been presented and these agents were thought to be probably simian in origin
- What diseases, if any, they might be responsible for in monkeys had not been established
- In other words, the researchers could not produce disease with these “agents” found in both inoculated and uninoculated cultures based upon identification by CPE and thus they were not “viruses”
- Many of these agents have been recovered from normal or uninoculated control cultures
- Hull et al. reiterate that eight apparently new submicroscopic, filterable, cytopathogenic agents recovered in tissue cultures of rhesus and cynomolgus monkey kidney tissue were described
- No definite associations were made between any of these agents and naturally occurring diseases of monkeys or other animals

- Dr. Stefan Lanka performed his own cell culture control experiments exposing the CPE fraud
- In control #1, normal cells were cultured with normal nutrient medium and only a small amount of antibiotics and on neither day one nor day five was any CPE found; the cells continued their normal, healthy growth
- In control #2, normal cells were again grown on normal nutrient medium and a small amount of antibiotics, but this time, 10% fetal calf serum was added to enrich the medium and still, the cells in the culture grew normally, both on day one and day five
- Control #3 included changing the nutrient medium to “minimal nutrient medium”—meaning lowering the percentage of fetal calf serum from the usual 10% to 1%, which lowers the nutrients available for the cells to grow, thereby stressing them—and tripling the antibiotic concentration
- On day five of the experiment, the characteristic CPE occurred, “proving” the existence and pathogenicity of the “virus”—except, at no point was a pathogenic “virus” added to the culture
- This outcome can only mean that the CPE was a result of the way the culture experiment was done and not from any “virus”
- Control #4 is the same control # 3 except that to this culture, a solution of pure RNA from yeast was added which produced the same result as column three, again proving that it is the culture technique—and not a “virus”—that is causing the CPE
- According to a cell culture technical bulletin, when cultures self-destruct for no apparent reason and no evidence of common biological contaminants can be found, cryptic “viruses” are often blamed
- It was stated that this is unfortunate, since the real cause of this culture destruction may be something else, possibly mycoplasma or a chemical contaminant, and as a result will go undetected to become a more serious problem
- The other factors which are said to be able to produce the CPE include:
- Bacteria
- Amoeba
- Parasites
- Exosomes
- Antibiotics
- Antifungals
- Chemical contaminants
- Age of the cell
- Environmental stress
- Cell culture experiments are supposed to include detailed mock-infections yet rarely do
- Mock-infections are supposed to be treated the same way as the experimental culture except without the “virus”
- Sometimes a “non-virulent strain” is used in the mock-infected specimen
- In a pivotal “SARS-COV-2” study by Zhou et al., the antibiotics in the experimental culture were doubled during the culture experiments to achieve a cytopathogenic effect in only 1 out of 24 cultures and this information was left out of the published study
- Regardless, mock-infections are not proper controls as only the variable being studied (the “virus”) should be eliminated
- As the unpurified fluids used also contain other host and foreign materials, contaminants, and pollutants, many confounding variables are eliminated as well if no human sample is used with the mock-infected culture
- Thus, a proper control for these pseudoscientific cell culture experiments would be utilizing a sample from a healthy host treated exactly the same way as the sample from the unhealthy host, “viral” transport media and all

If virology wants to be seen as a true science, it must adhere to the scientific method. This means that the experiments and the controls championed as the “gold standard” evidence for the field must be designed in adherence to this method. However, virology has it foundations rooted squarely in pseudoscience as there was never any naturally observed phenomena for which virology was built upon. The “virus” is an imaginary construct dreamt up in the minds of researchers who regularly failed to find a bacterial cause of disease who then assumed that there must be something else smaller and impossible to see contained within the fluids of sick humans. Like the very best Bigfoot hunters, virologists searched for these mythological entities for decades and failed to find direct evidence of them either directly within the fluids of a sick host or through various grotesque tissue cultures and inhumane animal experiments. The attempts to try and prove the existence of the fictional creations was on a downward trajectory, and by the 1950s, all was lost, and the field was ready to die its natural death.
However, things took a turn for the better for virology when John Franklin Enders introduced his fraudulent cell culture method in 1954 and ultimately revitalized the dying field. Virologists were now able to kill cells by combining various toxins in a Petri dish in order to create the picturesque works of art plastered all over the media as the representation of these mythological beings. The subsequent evidence and killing of the cell, referred to as the cytopathogenic effect, was redefined as a “virus” specific effect in order to establish an invalid dependent variable for the non-existent independent variable. Instead of direct evidence for the existence of these entities, a new form of indirect evidence was established in order to fool the masses with. Virology had successfully conjured up its own lab-created variables to play with rather than proving cause and effect through the study of any real-world phenomena.
While the field clung to the cell culture method in order to generate the pseudoscientific evidence needed to keep virology on life-support, there was a major problem that immediately popped up, as pointed out by Enders, the inventor of the technique, which he described witnessing during his initial experiments. The CPE blamed on the “virus” was observed during both the “infected” cultures as well as in his “uninfected” control cultures. This should have been the death knell for virology as it was clear that the toxic experimental design created the effect being blamed on the invisible “virus” as shown by the controls. If this wasn’t damaging enough, various other researchers throughout the remainder of the 1950’s reported the same findings as the identical CPE blamed on the “virus” was observed in healthy monkey control cultures said to contain no “viruses” whatsoever. Some researchers admitted that this effect could not be used to claim a “virus” was present and then relied on equally fraudulent antibody results to support their “virus” claims instead. Others attempted to state that the healthy control cultures contained “virus-like” agents yet failed miserably in attempts to prove pathogenicity using these fluids, thus showing instead that nothing meeting the definition of a “virus” was present. On top of this, many other factors became known to cause this same cytopathogenic effect other than the assumed “viruses” such as bacteria, amoeba, the antibiotics used, the age of the cell, etc. Virology had successfully refuted itself.
As the detailed descriptions of the control cultures proved the “virus” was not to blame for the observed CPE, many subsequent papers either stopped detailing what was done to the controls or simply avoided using controls all together. Controls known as mock-infections, if present in studies today, are ill-defined and have been shown in cases to be completely fraudulent, such as in the Zhou et al. paper, which treated the mock-infected control differently than the experimental culture, thus invalidating the results of a seminal “SARS-COV-2” paper. While these mock-infections are a form of control, they can hardly be considered proper controls as they do not eliminate only the “virus” particles from the equation. Also eliminated are the rest of the potential organisms, contaminants, chemicals, etc. within the fluids, which have been added to the “viral” sample that are subsequently left out of the mock-infected cultures, which contain no fluids from humans whatsoever. Thus, it can not be said that mock-infected cultures only change up the one variable being studied as there are many variables removed from the equation. Therefore, mock-infections without a comparable human sample minus the assumed “virus” is not a proper control.
If virology wants to be a true science, it must go back to the drawing board. Virologists must attempt to observe a natural phenoma where the independent variable (i.e. the cause; “virus”) can be observed in nature. At the very least, this means that they must find the particles that they believe are “viruses” directly in the fluids of a sick host and determine a way to get these particles away from everything else within the fluids. Virologists must determine a valid dependent variable (i.e. the effect being studied; symptoms of disease) in order to establish a testable and falsifiable hypothesis. Only then would a valid experiment be able to be designed, including the establishment of proper controls. Nowhere was this process carried out in order for Enders to create the cell culture method. As the cell culture is not a valid scientific experiment adhering to this process, any and all controls are scientifically invalid as a result. Thus, it really is a waste of time arguing whether or not virology uses controls as their methods are built upon a pseudoscientific foundation. We can argue for virologists to implement what would be proper controls given the pseudoscientific parameters of their experiment, but it is clear that even despite their lack of use of proper controls, virology disproved itself long ago.
It is time to focus instead on the heart of the issue, the lack of adherence to the scientific method. The best control of all is to have virologists walk us through how their field adheres to the scientific method. What natural phenomena was observed? How did the virologists identify and establish a valid IV? What is the DV? What hypothesis led to the creation of the cell culture method, and how was this determined? How was this cell culture method validated without first having successfully purified and isolated the “virus” in order to learn how to grow one? How can virologists justify using a lab-created effect to claim a cause that can not be observed until after the experiment takes place when this is the antithesis of the scientific method? Asking virology to show how it adheres to the scientific method is a control that they will fail at every time. It is the only control needed.

109 comments