
mock-infected
https://www.genscript.com/biology-glossary/10558/mock-infected#:~:text=A%20control%20used%20in%20infection,in%20the%20mock%2Dinfected%20specimen.
A control used in infection experiments. Two specimens are used one that is infected with the virus/vector of interest and the other is treated the same way except without the virus. Sometimes a non-virulent strain is used in the mock-infected specimen.”
A control in science is an element that remains unchanged or the same throughout the experiment. They allow for the experimentor to minimize the variables to the one that is being tested. There can be more than one kind of control used to ensure that all variables are accounted for. With the cell culture experiments performed in virology, the control culture, often referred to as the mock infected culture, is supposed to be treated in the same manner as the infected culture yet without the “virus” material present. This is supposed to show that the “virus” is the only variable that could be the cause of whatever effect is being studied and observed.
The problem for virology is that they do not have a proper independent variable (i.e. purified and isolated particles) to use in order to determine cause and effect. There are numerous substances said to be contained within the sample taken from a sick host including host materials, bacteria, fungus, multivesicular bodies, exosomes, other “viruses,” etc. The sample is always added to “viral” transport media which itself contains antibiotics, antifungals, fetal bovine serum, and other nutrients/chemicals. This sample is then added to an abnormal cell, usually from an animal such as an African green monkey, or from a cancer cell of some sort. The chosen cell has itself been cultured while out of its natural environment in unnatural media for an extended period of time. Before the culture experiments have even begun, there are numerous variables that can potentially be the cause of any observed effect created through experimentation. Thus, it is very important for virologists to perform proper controls to ensure that the results are valid and are in fact caused by their independent variable rather than a creation from their own experimental methods.
Unfortunately, this is not the case in virology. What is most often found in these papers, if found at all, is the mock-infected culture, which again is where they will take whatever cell line is chosen for the experiment and apply the same(?) compounds to it without the unpurified human sample. Sadly, it is often not clear exactly how the mock-infected culture was treated or if one was ever performed. However, even if done as described, is this a proper control which factors in all potential variables? No, absolutely not. The only way it would possibly be a proper control is if the virologists started with only the purified/isolated particles assumed to be the “virus” as the independent variable used for experimentation. As they do not do so, the unpurified human sample already has many host and foreign materials present along with many others added to it which need to be factored into the equation as potential causes of any observed effect. Thus, using only a cell without a human sample is not a proper control when an unpurified sample without isolated particles is utilized to try and prove cause and effect. In order to have a proper control, the virologists would need to use samples from both healthy humans and those who are sick with similar symptoms but said not to have the assumed “virus.” Ex-virologist Stefan Lanka summed this up brilliantly in his seminal paper The Misconception called Virus:
Virologists believe in viruses, because they add to the tissue and cell culture allegedly infected blood, saliva or other body fluids-after having withdrawn the nutrients from the respective cell culture and after having started poisoning it with toxic antibiotics. They believe that the cell culture is then killed by viruses. However, the death of the tissue and cells takes place in the exact same manner when no “infected” genetic material is added at all. The virologists have apparently not noticed this fact. According to the scientific logic and the rules of scientific conduct, control experiments should have been carried out. In order to confirm the newly discovered method of so-called “virus propagation”, in order to see whether it was not the method itself causing or falsifying the result, the scientists would have had to perform additional experiments, called negative control experiments, in which they would add sterile substances or substances from healthy people and animals to the cell culture.
These control experiments have never been carried out by the official “science” to this day. During the measles virus trial, I commissioned an independent laboratory to perform this control experiment and the result was that the tissues and cells die due to the laboratory conditions in the exact same way as when they come into contact with allegedly “infected”
material.
The purpose of control experiments is to exclude the posibility that it is the applied method or technique which may cause the result. Control experiments are the highest duty in science and also the exclusive basis of claiming that one’s conclusion is scientific. During the measles virus trial it was the legally appointed expert who stated that the papers which are crucial for the entire virology contain no control experiments. We learn from this that the respective scientists work extremely unscientifically, without noticing it.”
-Ex Virologist Dr. Stefan Lanka
https://archive.org/download/dr-stefan-lanka-the-misconception-called-virus/Dr%20Stefan%20Lanka%20-%20The%20Misconception%20Called%20Virus%20%28Jan%202020%29_text.pdf
What would happen if virologists actually performed proper controls? What would they observe happening in their petri dishes? What kind of particles would they find upon electron microscopy examination of their cultured creations? While virologists may not want to perform the proper controls necessary to validate their own findings, we have a few sources where some form of control was carried out. What they showed is that, when these controls are performed, the entire cell culture process and virology itself is completely invalidated as the same particles and effects are observed even in the absence of “viral” material.
Control # 1: John Franklin Enders 1954 Measles Paper
In 1954, John Franklin Enders ushered in the era of the cell culture experiments in virology after virologists repeatedly failed to isolate their filterable “viruses” (translated from German):
However, these experiments failed to produce growth of the filterable virus. In the hope that various animal experiments have also been carried out to trigger virulence of filter-passing virus carried out. But the results were always negative. It never succeeded from the filtrates by renewed inoculation on the various culture substrates a filterable microbe (“a true filter-passing virus”).” – Prof. Karlheinz Lüdtke, Max-Planck-Institut für Wissenschaftsgeschichte, Frühgeschichte der Virologie, Sonderdruck 125, 89 Seiten, 1999. i. K. (A 2)
https://impfen-nein-danke.de/u/P125.pdf
Enders cell culture technique established in his 1954 measles paper is essentially the exact same process carried out today in order to “isolate” any “virus.” It consisted of culturing the human sample along with a mixture of many added chemicals and foreign substances.
Interestingly, Enders did do a version of the control procedure other virologists are supposed to carry out when performing these kinds of experiments. In an uninoculated monkey kidney cell cultured in the same way as the sample supposedly containing the measles “virus,” Enders observed the exact same cytopathogenic effect in the cell culture that he and others attributed to “viruses:”
“Monkey kidney cultures may, therefore, be applied to the study of these agents in the same manner as cultures of human kidney. In so doing, however, it must be borne in mind that cytopathic effects which superficially resemble those resulting from infection by the measles agents may possibly be induced by other viral agents present in the monkey kidney tissue (cf. last paragraph under G) or by unknown factors.”
“A second agent was obtained from an uninoculated culture of monkey kidney cells. The cytopathic changes it induced in the unstained preparations could not be distinguished with confidence from the viruses isolated from measles. But, when the cells from infected cultures were fixed and stained, their effect could be easily distinguished since the internuclear changes typical of the measles agents were not observed. Moreover, as we have already indicated, fluids from cultures infected with the agent failed to fix complement in the presence of convalescent measles serum. Obviously the possibility of encountering such agents in studies with measles should be constantly kept in mind.”
Sadly, even though Enders found that the same effect can be observed without “virus” material present in the culture, his confirmation bias led him to conclude that there must have been some other “virus” or agent in his culture which lead to the death of the cell rather than occurring as a result of the cell culture procedure itself. Fortunately, he was not the only one to observe this phenomena in uninoculated cultures:

Control # 2: Stefan Lanka’s Phase One Controls
Recently, Dr. Stefan Lanka himself has taken on the challenge of performing the cell culture controls that send virologists cowering in fear. In essence, he performed the exact same cell culture steps without the added “viral” material and was able to produce the exact same cytopathogenic effect Enders and other’s noticed in their uninoculated cultures by manipulating the additives, thus disproving this effect as due to any “virus.” You can find a good breakdown of the methods utilized here with an English translation:
Kontrollexperiment Phase 1 – Mehrere Labore bestätigen die Widerlegung der Virologie durch den cytopathischen Effekt
If you would like to hear Dr. Lanka speak about the controls in a short 7 min clip, you can view that here:
However, my favorite explanation comes from Dr. Tom Cowan’s excellent booklet Breaking The Spell:
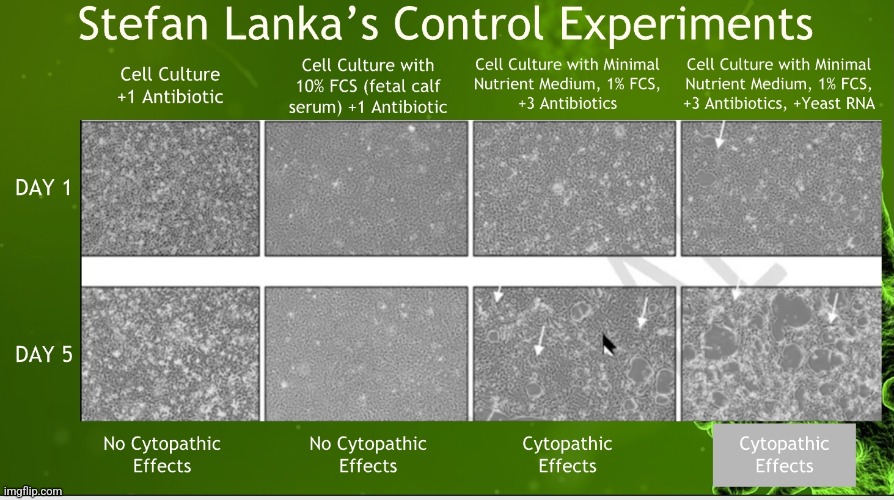
“Here is the essence of Lanka’s experiment, done by an independent professional laboratory that specializes in cell culturing. As seen in this series of photographs, each of the four vertical columns is a separate experiment. The top photo in each column was taken on day one, and the bottom photo was taken on day five.
In vertical column one, normal cells were cultured with normal nutrient medium and only a small amount of antibiotics. As you can see, on neither day one nor day five was any CPE found; the cells continued their normal, healthy growth.
In vertical column two, normal cells were again grown on normal nutrient medium and a small amount of antibiotics, but this time, 10% fetal calf serum was added to enrich the medium. Still, the cells in the culture grew normally, both on day one and day five.
The third vertical column shows what happened when Dr. Lanka’s group used the same procedures that have been used in every modern isolation experiment of every pathogenic virus that I have seen. This included changing the nutrient medium to “minimal nutrient medium”—meaning lowering the percentage of fetal calf serum from the usual 10% to 1%, which lowers the nutrients available for the cells to grow, thereby stressing them—and tripling the antibiotic concentration. As you can see, on day five of the experiment, the characteristic CPE occurred, “proving” the existence and pathogenicity of the virus—except, at no point was a pathogenic virus added to the culture. This outcome can only mean that the CPE was a result of the way the culture experiment was done and not from any virus.
The fourth and final vertical column is the same as vertical column three, except that to this culture, a solution of pure RNA from yeast was added. This produced the same result as column three, again proving that it is the culture technique—and not a virus—that is causing the CPE.”
Control # 3: Exosome Research
In the early 1980’s, researchers working with cell cultures of reticulocytes (slightly immature red blood cells) found small 40-150 nm vesicles in their electron microscopy images. Even though they were not working with “viral” material in their cultures, the researchers found particles that were the same size, shape, and density as “viral” particles. While the particles were originally thought to be cellular debris and/or garbage bags, they were assigned a theoretical function related to intercellular communication. Even though the vesicles were indistinguishable from “viral” particles, they were eventually given the name exosomes:
“30 years ago, a paper in JCB (Harding, Heuser and Stahl, 1983) and one in Cell (Pan and Johnstone, 1983)—published within a week of each other—reported that, in reticulocytes, transferrin receptors associated with small ∼50 nM vesicles are literally jettisoned from maturing blood reticulocytes into the extracellular space. The name “exosome” for these extracellular vesicles was coined a few years later by Rose Johnstone, although the term had in fact been used a few years earlier, when referring to other membrane fragments isolated from biological fluids (Trams et al., 1981; the term “exosome complex” has also been used for a totally different entity: namely, the intracellular particle involved in RNA editing [Mitchell et al., 1997]).”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3575527/#!po=0.909091
What has occurred since the discovery of these “virus-like” particles is a blurring of the lines between two sets of disciplines (exosome/virology) using the same particles as a representation for their chosen fictional entity. As the decades progressed, exosomes have become even more “virus-like,” to the point that they are now said to carry “viral” proteins and materials and can infect cells:
“Virus-infected cells release exosomes that are implicated in infection through transferring viral components such as viral-derived miRNAs and proteins. As well, exosomes contain receptors for viruses that make recipient cells susceptible to virus entry.”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7293471/
Going through the major exosome papers of the 1980’s shows that the exact same cell culture procedures and experiments were being done without “viral” material present yet the same exact “virus-like” particles as seen in virology were still being found. In the early research papers, materials from animals were mostly used. However, eventually samples from healthy humans were cultured and shown to contain these “virus-like” particles. In other words, the exosome researchers were carrying out the control cultures which showed that the methods produced the results as no “virus” was necessary.
In this pivotal exosome study from 1983, sheep reticulocytes were cultured in various ways (the process was outlined in previous papers). They were incubated for 36 hours and eventually “separated” from the culture medium. The vesicles with “surface knobs” that were observed looked very reminiscent of a certain group of “coronaviruses:”
Fate of the transferrin receptor during maturation of sheep reticulocytes in vitro: selective externalization of the receptor

“Using sheep reticulocytes, surface-labeled with ‘251, we observed that the amount of ‘251-labeled plasma membrane protein precipitated with the specific anti-transferrin-receptor antibody decreases with time in culture in parallel with the loss of reticulum (Pan, Blostein, and Johnstone, 1983). These data suggested that the receptor might be removed from the plasma membrane. To test this possibility the cell surface was labeled with ‘251 and lactoperoxidase (Reichstein and Blostein, 1975) and the cells were incubated for 36 hr at 37°C. Initially, and after 36 hr, the cells were separated from the incubation medium. Plasma membranes were prepared (Dodge et al., 1963) and solubilized, and the solubilized membranes and cell-free postincubation medium were passed through an anti-transferrin-receptor immunoaffinity column as described (Pan, Blostein, and Johnstone, 1983).”
doi: 10.1016/0092-8674(83)90040-5.
In this study from 1985, once again sheep reticulocytes were cultured in various ways previously outlined over many prior studies. It showed that long-term incubation in culture created the smaller particles observed in electron microscopy images. It can be seen in the bottom image description that 1% bovine serum albumin and PBS were added as well as fresh culture medium before imaging. The process observed in the EM image is said to be exocytosis, which has been claimed to be the particles emerging from the cell to transport information. However, is exocytosis being observed or could it be “viral” particles entering the cell through endocytosis? The interpretation is in the eye of the beholder:
Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes

“In the present study using an immunological approach, we have obtained visual evidence that the MVEs are the penultimate structures from which the transferrin receptor appears to be released during long-term incubation of sheep reticulocytes. The data show the changing nature of the intracellular structures that contain transferrin receptors during long-term incubation and that ultimately, small bodies with the transferrin receptor at the external surface are released.”

doi: 10.1083/jcb.101.3.942.
In this 1986 study, it is admitted that many different types of cultures release these exact same particles into the culture medium. These consisted of cultured brain cells including C6 glioma and two cell lines of mouse neuroblastoma, B16 mouse melanoma cells, and calcifying cartilage in the long bone of fetal calves where matrix vesicles can be released by collagenase treatment. The researchers in this study cultured rat glioma cells which were obtained at passage 39, HeLa cells cultured with 5 mM sodium butyrate, and Friend erythroleukemia cells grown in suspension. You can see that similar cell culture techniques used in virology were utilized in this paper to create the same particles:
Exfoliation of the/3-adrenergic receptor and the regulatory components of adenylate cyclase by cultured rat glioma C6 cell
“It has recently been observed that some cultured cell lines release packets of material into the medium that appear to be derived from the plasma membrane. Trams et al. [1] found that several cultured brain cells including C6 glioma and two cell lines of mouse neuroblastoma release particles of 500-1000 nm in diameter which contain 5′- nucleotidase and a divalent cation-activated ATPase. They also observed the release of cell-surface 5′-nucleotidase into the culture medium from astroblast, fibroblast and melanoma cell lines.
Others had previously demonstrated that B16 mouse melanoma cells shed plasma membrane-derived vesicles into the medium [2]. A similar phenomenon occurs in calcifying cartilage in the long bone of fetal calves where matrix vesicles can be released by collagenase treatment [3].”
“Cell Culture. Rat glioma cells were obtained at passage 39 from the American Type Culture Collection (Rockville, Maryland) and grown as described previously [12,14]. HeLa cells, strain 30002 [15], were cultured with 5 mM sodium butyrate [15,16] and treated with (-)-isoproterenol to down-regulate their B-adrenergic receptor [17]. Friend erythroleukemia cells were grown in suspension as previously described [16].”
Preparation of Exosomes and Membranes. For preparation of exosomes, C6 cells were grown to confluency in 530 cm z NUNC dishes (A/S NUNC, Rochilde, Denmark). 24 h prior to exosome collection, the growth medium was removed and monolayers were rinsed four times over a period of 1.5 h with serum-free medium adjusted to contain 5 mM MgC12 and 2 mM CaCI 2- The cells were cultured in the same medium for 22.5 h. The medium was then collected and centrifuged at 10000 × g for 30 min at 4°C. The supernatant was carefully removed and centrifuged at 65,000 rpm for 90 min using a Beckman Ti 70 rotor. The pellets (exosomes) were washed once with 10 mM Tris-HC1 (pH 7.4) resuspended and homogenized “in the same buffer (0.5 mg of protein per ml) and stored for further analysis in liquid nitrogen. Crude membranes from the different cell lines were prepared as previously described [18].”
doi: 10.1016/0167-4889(86)90184-9.
In this 1987 study, we can see that the same exact compounds used in virology were utilized for the creation of exosomes: Eagle’s minimal essential medium supplemented with glutamine (4 mM), adenosine (5 mM), inosine (10 mM), penicillin (200 units/ml), and streptomycin (200 pg/ml) (standard culture medium). While sheep reticulocytes were used for initial culturing and study, the researchers wanted to see if they could find these same particles by culturing mature erythrocytes as well as washed, unfractionated blood cells from phlebotomized sheep and enriched fractions of white cells. Sure enough, they were able to create and find the exact same particles in numerous cultures:
Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes)
“Reticulocytes were prepared as previously described from phlebotomized sheep (6, 11). Cells were cultured for 24-40 h at 37 “C in roller bottles in Eagle’s minimal essential medium supplemented with glutamine (4 mM), adenosine (5 mM), inosine (10 mM), penicillin (200 units/ml), and streptomycin (200 pg/ml) (standard culture medium). Cells were collected by centrifugation at 8,000 X g for 10 min. The cell-free supernatant was recentrifuged for 90 min at 100,000 x g to collect the released vesicles. To assess whether cells other than reticulocytes gave rise to vesicles, washed, mature erythrocytes as well as washed, unfractionated blood cells from phlebotomized sheep and enriched fractions of white cells were cultured. Putative vesicle fractions were collected from all cultures after removal of the cells as described above. The pelleted vesicles were stored at -70 “C until required or used immediately after preparation.
Plasma membranes from reticulocytes and mature cells were prepared by osmotic lysis in 5 mM phosphate buffer, pH 8.0, containing 1 mM EDTA as described (15). The membranes were washed and stored at -70 ‘C in 20 mM phosphate buffer, pH 7.0. The membranes were centrifuged before use and resuspended in the appropriate medium as given in the text.”
https://pubmed.ncbi.nlm.nih.gov/3597417/
As can be seen from the above studies, the particles assumed as exosomes were created by the same cell culture process utilized by virologists with materials from healthy animals. No “viral” sample was necessary in order to create the same exact EM images as seen by virologists. I previously discussed how this exosome concept, with an origin tied to the cell culture tricks used in virology, was created. As the same “viral” particles were found in healthy subjects, these results obviously could not stand as it would destroy the very foundation that virology was built upon. Thus, the “virus-like” particles were given a different name and function in order to keep the “virus” lie intact. The exosome became the escape clause for researchers to fall back on so that virology could continue marching ahead unscathed by these contradictory findings.
In fact, it wasn’t until the early 2000’s when exosomes were finally said to be found outside of the cell culturing process. The exosome results up to at least 2005 were only applicable to experiments done in vitro (i.e. outside the living body and in an artificial environment). There was no evidence that these particles existed without the culture process:
“Many cell types release exosomes in vitro but there is still little evidence of whether exosomes are produced in vivo.”
Exosomes are small membrane vesicles mostly secreted by hematopoietic cells in culture medium. In the present work, we show that a population of vesicles sharing characteristics with exosomes can be isolated from the plasma of healthy donors. Differential ultracentrifugations of blood plasma allowed us to purify vesicles similar in size and shape to the previously described exosomes (reviewed in 3). It clearly appeared that blood plasma is very rich in various contaminants from diverse origins.”
doi: 10.1093/intimm/dxh267.
It should be clear from these early exosome experiments why this fictitious entity is so closely entwined with the “virus.” It has been repeatedly shown that the same particles claimed to be “viruses” can be found in the supernatant made from cultures containing no “viral” material whatsoever. It has been stated repeatedly that “viruses,” exosomes, and similar particles can not be separated from each other:
“Exosomes have been the most studied in the context of infection. An important note, however, is that exosome purity was not always analyzed in these studies, and therefore, the vesicle population may have consisted of both exosomes and microvesicles, which overlap in size and density.”
https://www.embopress.org/doi/full/10.15252/embr.201439363
“The fact that exosomes are similar in size, shape, and density to many viruses makes separating exosomes from viruses a veritable challenge.”
https://www.frontiersin.org/articles/10.3389/fcimb.2021.671625/full
This is because the very same particles are being picked as a representation by different researchers as different entities in every single case. It is a fact that exosomes have their origin in the same culture process used by virologists, have the same size/shape/density as “viruses,” and are claimed to have the exact same “viral” proteins/materials as well the ability to hijack cells to create “viruses.” Exosomes are “viruses” and “viruses” are exosomes. Both are fictional entities/concepts used to explain lab-created results in a petri dish. However, if you are not convinced that we are dealing with the same particles created through the same process, maybe these next pieces of information will provide further evidence that it is the method which creates the results seen and not the particles observed afterwards.
In 2019, a study came out claiming that exosomes can, in fact, produce the exact same cytopathogenic effect (i.e. structural changes in host cells that are caused by “viral” invasion) said to be SPECIFIC to “viruses.” While I have gone over CPE before and shown that it can be caused by many factors related to the culture conditions, virologists still use it as the evidence a “virus” is present in the sample. Granted, sometimes they will claim a “virus” is present even without observing CPE but that is another article entirely. In any case, exosomes can now be added to the long list of factors other than a “virus” which can induce this effect, thus blurring the line between these “distinct entities” even further. And once again, we can see the exact same “virus-like” particles were found in both the “uninfected” and the “infected” cell culture EM images:
Exosomes Carry microRNAs into Neighboring Cells to Promote Diffusive Infection of Newcastle Disease Virus

Exosomes Isolated from NDV-Infected HeLa Cells Cause a Cytopathic Effect
“Several studies have shown that exosomes are involved in virus infections. However, the effect that exosomes have on NDV-infected tumor cells is not known. In this study, we focus on the role of exosomes secreted by NDV-infected HeLa cells in promoting NDV replication. Three miRNA candidates (miR-1273f, miR-1184, and miR-198) embraced by exosomes were associated with enhancing NDV-induced cytopathic effects on HeLa cells.”
“These results confirm that the pellets from NDV-infected and naive HeLa cells were exosomes, and thus, these samples were prepared for further study. HeLa cells were incubated with these particles for 24 h and then infected with NDV to observe the cytopathic effects. To our surprise, the cytopathic effect was significantly observed in cells treated with exosomes from NDV-infected cells (Figure 2D).”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6631457/
In October 2019, another study looking at exosomes released from Acanthamoeba castellanii also found that the exosomes induced a cytopathogenic effect in the cells:
Quantitative proteomic analysis and functional characterization of Acanthamoeba castellanii exosome-like vesicles

“To examine whether EVs of A. castellanii also lead to cytotoxic effects on C6 cells, the CPE of C6 cells was evaluated after co-culture with EVs from A. castellanii for 24 h. The data showed that C6 cells markedly detached from the culture plates after coincubation with EVs from A. castellanii compared with the PBS-treated control, suggesting that parasite-derived EVs destroyed the adhesive ability of target cells and caused cell death (Fig. 3a). Time-lapse microscopy analysis also revealed that the C6 cells were more spherical and cytolytic following exposure to EVs from A. castellanii (Fig. 3b).”
“It has been demonstrated that A. castellanii EVs display cytotoxicity to Chinese hamster ovary (CHO) and T98G mammalian cells via necrosis and apoptosis, respectively, suggesting that different cell death mechanisms are induced by EVs in various host cells [33, 35]. We also showed that EVs cause cell disruption and reduce cell adhesion ability, resulting in CPE on rat glial C6 cells, which is similar to the outcome exerted by the secreted proteins [15].”
https://link.springer.com/article/10.1186/s13071-019-3725-z
In Summary:
- A control in science is an element that remains unchanged or the same throughout the experiment and it allows for the experimentor to control the variables
- Virology sometimes uses mock-infected cultures where two specimens are used: one that is infected with the “virus/vector” of interest and the other is treated the same way except without the “virus”
- According to ex-virologist Stefan Lanka:
- The death of the tissue and cells takes place in the exact same manner when no “infected” genetic material is added at all
- The scientists would have had to perform additional experiments, called negative control experiments, in which they would add sterile substances or substances from healthy people and animals to the cell culture
- These control experiments have never been carried out by the official “science” to this day
- The purpose of control experiments is to exclude the posibility that it is the applied method or technique which may cause the result
- Control experiments are the highest duty in science and also the exclusive basis of claiming that one’s conclusion is scientific
- Control # 1: In 1954, John Franklin Enders, creator of the cell culture technique still used by virologists today, disproved his own methods:
- Cytopathic effects which superficially resemble those resulting from infection by the measles agents may possibly be induced by other “viral” agents present in the monkey kidney tissue or by unknown factors
- A second agent was obtained from an uninoculated culture of monkey kidney cells
- The cytopathic changes it induced in the unstained preparations could not be distinguished with confidence from the “viruses” isolated from measles
- Control #2: Lanka’s experiment, done by an independent professional laboratory that specializes in cell culturing, consisted of four controls:
- In control 1, normal cells were cultured with normal nutrient medium and only a small amount of antibiotics – no CPE by day 5
- In control 2, normal cells were again grown on normal nutrient medium and a small amount of antibiotics, but this time, 10% fetal calf serum was added to enrich the medium – no CPE by day 5
- In control 3, this included changing the nutrient medium to “minimal nutrient medium”—meaning lowering the percentage of fetal calf serum from the usual 10% to 1%, which lowers the nutrients available for the cells to grow, thereby stressing them—and tripling the antibiotic concentration – CPE was observed by day 5
- In control 4, yeast RNA was added and this produced the same result as control 3, again proving that it is the culture technique—and not a “virus”—that is causing the CPE
- Control # 3: In the early 1980’s, researchers working with cell cultures of reticulocytes (slightly immature red blood cells) found small 40-150 nm “viral-like” particles in their electron microscopy images without “viral” material present
- In 1983, “coronavirus-like” particles were found in cell cultures of sheep reticulocytes
- In 1985, the same particles were seen after long incubation periods in EM images
- In 1986, numerous cell cultures were admitted to produce the same particles:
- Cultured brain cells including C6 glioma and two cell lines of mouse neuroblastoma
- B16 mouse melanoma cells
- Calcifying cartilage in the long bone of fetal calves where matrix vesicles can be released by collagenase treatment
- Rat glioma cells which were obtained at passage 39
- HeLa cells cultured with 5 mM sodium butyrate
- Friend erythroleukemia cells grown in suspension
- In 1987, the same particles were seen in mature erythrocytes as well as washed, unfractionated blood cells from phlebotomized sheep and enriched fractions of white cells and the same culture ingredients were described:
- Eagle’s minimal essential medium supplemented with glutamine
- Adenosine
- Inosine
- Penicillin
- Streptomycin
- In a 2005 study, it was admitted that many cell types release exosomes in vitro (in the lab) but there was still little evidence of whether exosomes are produced in vivo (in the body)
- Exosomes are small membrane vesicles mostly secreted by hematopoietic cells in culture medium
- It clearly appeared that blood plasma is very rich in various contaminants from diverse origins
- Exosomes have been the most studied in the context of infection, however, exosome purity was not always analyzed in these studies, and therefore, the vesicle population may have consisted of both exosomes and microvesicles which overlap in size and density
- The fact that exosomes are similar in size, shape, and density to many “viruses” makes separating exosomes from “viruses” a veritable challenge
- In a June 2019 study, exosomes isolated from NDV-infected HeLa cells cause a cytopathic effect
- Several studies have shown that exosomes are involved in “virus” infections
- Three miRNA candidates (miR-1273f, miR-1184, and miR-198) embraced by exosomes were associated with enhancing NDV-induced cytopathic effects on HeLa cells the cytopathic effect was significantly observed in cells treated with exosomes from NDV-infected cells
- In a October 2019 study, the researchers examined whether EVs of A. castellanii also lead to cytotoxic effects on C6 cells, the CPE of C6 cells was evaluated after co-culture with EVs from A. castellanii for 24 h
- The data showed that C6 cells markedly detached from the culture plates after coincubation with EVs from A. castellanii compared with the PBS-treated control, suggesting that parasite-derived EVs destroyed the adhesive ability of target cells and caused cell death
- The researchers showed that EVs cause cell disruption and reduce cell adhesion ability, resulting in CPE on rat glial C6 cells

Proper controls are essential in order to ensure that the results obtained in a scientific experiment were actually caused by the variable being studied and were not a creation stemming from the methods employed. If virologists were to carry out proper controls on their cell culture experiments, they would easily see that the very methods they use are creating the effects they want to observe. Fortunately, there are instances where controls were performed by researchers both in and outside of virology which absolutely show that the exact same “virus” particles can be found in cell cultures without any “virus” material ever being present. From John Franklin Enders invalidating his own cell culture technique in his 1954 measles paper to the numerous examples of researchers finding the same “virus-like” particles in materials from healthy animals in exosome research, the evidence shows that it is the cell culture process, and not the “virus” material, which creates the desired effect.
Sadly, this realization was not taken up by the scientific community. Rather than admitting that the cell culture method and the conclusions drawn are flawed, a whole new off-shoot of research was created around a new theoretical entity in exosomes. As time has passed, exosomes have incorporated even more “viral” characteristics to the point that they are considered “non-infectious viruses.” A January 2019 study pointed out that it is essentially up to the researcher to decide which is which:
“Intermediate particles exist on the spectrum between virus and exosome that contain both host and viral components, making it nearly impossible to classify these vesicles as either defective viruses or exosomes that contain viral components [22]. Intermediate particles are often classified as a virus or exosome derivative, depending on the preference of the investigator, but once these vesicles deviate from strict definitions they may be more accurately defined as an assortment of lipid-encased particles that cannot be easily differentiated [22].”
https://www.microbiologyresearch.org/content/journal/jgv/10.1099/jgv.0.001193?crawler=true

What is clear is that the exosome is the perfect escape clause for virology. There is always some sort of rescue device available to the researchers to explain away contradictory results. The exosome researchers, performing the exact same cell culture process as done in virology, found the exact same particles in healthy material. The research completely disproved that “viral” particles are replicating in the petri dishes. Rather, the results showed that the cell culture method destroys the cell causing it to break apart into small pieces. The cellular debris, as exosomes were originally claimed to be, have been picked as representatives for many fictional entities including “viruses” and exosomes. The exosome research should have been the final nail in the coffin for virology. These experiments served as the controls virology is terrified of and will avoid performing. Instead, we have a whole new concept to explain away the contradictions.

Hi Mike! 😊. As you know I am taking a break from the front lines in this seemingly never-ending battle, but I decided to take a peek at your latest blog post since (as usual!) I could tell it was not only important but brilliant! Your work is only getting better and better and the nails in the coffin if virology are only getting tighter and tighter. The way you wrote this post is so clear, and the evidence is so devastating. Any last-gasp defenses put forth by “the other side” will seem weak indeed — but only to those who have their minds turned to the “on” position! It takes so much effort, both intellectually and emotionally, to disabuse oneself of these misteachings. But you are the brilliant presenter who makes it all much easier and clearer. Without your work, I daresay we would all be in a weaker position indeed. It’s so important to have clear thinkers and writers like you. So this is just a little love note saying thank you! I wish (sometimes) that I could still hang out on the front lines with you all, but I am taking care of my own “stuff” and honestly learning important lessons about how I do t always have to be going to battle. But that’s just me. The battle is highly important and I will keep checking back in to see how my favorite warriors are doing! Keep up the great work, I know you will! It’s in your blood (although I can’t prove that scientifically!) haha.
LikeLiked by 1 person
Hi Carolyn! As always, I definitely appreciate the kind words and support! I miss having your insight but I definitely understand the need to step back and take care of oneself. I have slowed down a little as these articles seem to take longer than they used to in order to research and write but my hope is that I present the information properly. Please keep in touch! 🙂
LikeLike
From the blog post:
“It is a fact that exosomes have their origin in the same culture process used by virologists”
That’s your false “originalist” pet theory again, as exposed by the ‘clean’ fluids papers Michael S posted.
Okay let’s keep building on the toroidal primordial soup that both originally begat metabolic evolution and *continues* to beget metabolic evolution every day.
The cytoplasm in every cell is that enduring primordial Soup at harbor. The cell membrane is an enduring, reliable, microscopically-built harbor within which the vast (‘electrochemical’) energetic oceans can be bestilled by the beating heart of consciousness, so that organic holograms/animations may exist as ‘carbon based lifeforms.’ (The Soup of course also exists in waters outside of cells and waters outside of organisms.) The human is a hyperevolved culture (conglomerate) of nested cytoplasmic harbors: inside the cell nucleus there are the cytoplasmic waters of the nucleoplasm distinctly nested within the larger cell cytoplasm.
The nested cytoplasmic Soup relates to the amoeba exosomes in this blog post being falsely implicated in ‘aiding and abetting’ a ‘viral infection’ which in reality is just what we might call the exosomal domino effect. We all know what the domino effect is. A positive feedback loop is one type of domino effect. In this case, one exosome, the one wrongly accused of being a “virus” and causing “NDV,” has entered a cell that is maladapted to the Terrain via a trauma of some kind, and has communicated to the nucleic acid formations in the primordial cytoplasmic soup in such a way as to cause an(other) exosome (microRNA this time) with a subset of the RNA of the first exosome to to come into being.
How does it come into being? Exactly like the ‘amplifications’ come into being in the PCR test – they’re born. In the cytoplasm are polymerases and free nucleic acids. The primary exosome (being blamed for ‘NDV’) is the equivalent of the template in the PCR test, right? In the PCR test the template is the supposed ‘virus’ that the synthetic primers attach to in order to catalyze the growing of the rest of the template onto the end of the primer, as grown with the free nucleic acids by the polymerase, in order to duplicate (amplify) the template.
The primer is presumably made in the inner sea harbor we call the nucleolus. When the primary exosome (the mRNA template) entered into the outer harbor (cytoplasm) it caused the maladapted, decentralized intelligent cell to need to amplify the message in order to alert the centralized body intelligence that it is having a hard time adjusting to the nature of the message coming from the surrounding environment, which it does by reproducing the primary exosome because if it is having trouble adjusting to the message and its neighbor cells also have trouble then that’s bigger trouble for the body than if it was just the one cell; therefore amplification is the only recourse for the maladapted cell in a ‘centralized’ system of intelligence. So inside the nucleolus the cell manufactures the relevant primers (microRNA) for the template and the primers move into the cytoplasm and one of them bonds with the template (primary exosome). The template is duplicated and is now DNA instead of RNA. Then primordial ‘transcription’ takes place and the DNA template’s peptide bonds break and it becomes two mRNA templates identical to the original, whereby two more primers (microRNA) that were made in the nucleolus do *their* thing along with more polymerase and free nucleic acids in order to ultimately turn the two mRNAs into four, then eight, and so on.
Evolution (adaptation) in organisms is the primordial Soup doing its thing inside themselves in symbiosis with the overlayed, organismal intelligence being responsive to local disequilibria. A fundamental dynamic of that primordialism is the proteomic polymerase chain reaction of intercellular communication.
The cell presumably sends out the secondary microRNA exosomes so that neighboring cells have pre-made primers for their amplifications; this is the return function for homeostatic purposes in the once-again toroidal dynamic – the in-built redundancy so as to avoid the ‘game of telephone’ whereby the message gets distorted. Therefore we can see why the ignorant germ theorists would associate the homeostatic microRNA secondary exosome with a disease causing mechanism in a cell culture that has no mother organism to be responsive to cell needs in the organic way that they need to be responded to.
LikeLike
“It is a fact that exosomes have their origin in the same culture process used by virologists”
That’s your false “originalist” pet theory again, as exposed by the ‘clean’ fluids papers Michael S posted.”
Michael S posted no such paper. It is a FACT exosomes were first seen as results of cell culture experiments. They were first “discovered” in culture in 1981. The other major studies elucidating on exosomes in the 1980’s and 90’s were from cell culture. You are the only one presenting false information here.
The rest of your comment is quite the fictional story with plenty of presumptions. Care to share any proof which follows the scientific method that shows exosomes exist and function as you claim?
LikeLiked by 1 person
I tried posting a comment here before, but it never showed up. That comment is here: https://jeffgreenhealth.substack.com/p/are-viruses-merely-cell-debris
Exosomes that contain some level of viral protein cannot be separated without destroying both proteins. This is the nature of such entities due to their scale. In hindsight, there is no need to do so since viruses and exosomes appear in separation much of the time and can be observed separately and their structures identified.
Exosomes are not merely cellular debris, nor are viruses, as you are suggesting. They have very clear differences in their structure, even in the pictures you are showing. Although the pictures you are showing are of low quality, there are many that exist that clearly show their differences in greater detail. The glycoproteins of each are quite different in structure and distinguish viruses, exosomes, and cell debris from each other. Further, adenoviruses, which are icosahedral in form, are entirely different in shape than exosomes and could never be confused for extracellular vesicles. The same is true for bacteriophages, a type of hybrid bacterial virus. You are using spherical viruses, like coronaviruses, which may at first appear as some form of vesicle.
On their surface, many cells and entities in the body appear to be similar or the same, until closely observed. The exosome is clearly a container of fluids that communicate and impart their fluids to cells. These entities must exist in the body to facilitate cohesive communication. You should agree that even on a theoretical level, they must exist. Viruses, however, are special protein structures manufactured and replicated (sometimes highly) by cells in distress that carry enzymatic glycoproteins on their capsid, operating on mRNA and/or DNA. They are entirely different than an exosome, which is not replicated, and they are especially not mere cellular debris.
LikeLiked by 1 person
Do you have any study which adheres to the scientific method showing completely purified/isolated particles assumed to be exosomes where they were able to be characterized and their functions proven?
If the exosome and “virus” particles are unable to be separated, not only would they not be able to be characterized properly, they would not be able to be distinguished from each other.
As “viruses” and exosomes have never been purified and isolated properly, one can not know what a “virus” looks like nor what an exosome looks like. One can also not know which proteins belong to which. There is no way to tell what is an exosome from a “virus” as they are the same cellular debris created from the same experimental processes.
LikeLiked by 1 person
You have likely already quoted from multiple studies that contain a method section that is replete with information on how particle(s) were isolated, so I will refrain from going on a hunt for a study you will likely not agree with in terms of isolation.
Functionality and isolation are two separate things. Functionality is the action an entity takes. In static cultures, the actions of those entities are not as they would appear in the body since static environments are devoid of the entirety of the microbiome and the body. However, particles can still be characterized.
The pictures you are providing in your article are isolated particles of exosomes and viruses, whether naturally existing or not. You seem to have a misunderstanding about what isolation is. You are seemingly claiming that viruses and exosomes do not exist because they have not been “completely isolated”, yet do not seem to understand that isolation standards allow for around a 1% contamination in their isolate when filtrated through a 0.22 µm pore size. It is a delicate process but normally does not hinder observation when properly conducted. In those circumstances, you can still characterize and identify those particles. And, it is obvious that isolation is taking place, or such pictures would not exist, to begin with.
You are taking the stance of Kaufman/Cowan/Lanka, and redefining isolation to conform to some impossible standards of “complete isolation” that cannot take place by the very nature of isolation itself. It is an impossible threshold to meet and is not founded on scientific reason or common sense.
It is not accurate whatsoever to claim you cannot tell these various particles apart. If this were true, the pictures of viruses, exosomes, and cellular debris would all look exactly the same—they do not.
LikeLiked by 1 person
You must not understand that the scientific method requires a valid independent variable that must be able to be manipulated and varied in order to prove cause and effect. Without completely separating the assumed “viral”/exosome particles from others that are the same size, shape, and density, they can not know which they are working with nor what proteins belong to which particle, if any. They also can not prove cause and effect without the independent variable as if the particles are not completely separated from everything else, there are other variables that could potentially be causing any effect observed. If they can not separate the particles, there is no way to know which particles in EM are the ones the researchers are looking for. As the particles can not be observed alive, their functions can not be proven.
I am not making the claim that they can not tell the particles apart nor separate them. The researchers are:
“Moreover, we emphasize that in the specific case of virus-infected cells, IT IS ALMOST IMPOSSIBLE TO DISTINGUISH EVs FROM (noninfectious) VIRUSES AND TO SEPARATE THEM.”
“Because EVs are produced by virtually all cells, probably every viral preparation is in fact a mixture of virions and EVs. To study their respective functions, IT IS NECESSARY TO SEPARATE EVs AND VIRIONS.”
“Here, we aimed to create awareness that virus preparations may never be pure but rather are contaminated with diverse subpopulations of EVs, and some of these EVs MAY BE EITHER INDISTINGUISHABLE FROM OR VERY SIMILAR TO SO-CALLED DEFECTIVE VIRUSES.”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4995926/
“HOW CAN WE BE SURE THAT WE ARE ISOLATING AND QUANTIFYING EXTRACELLULAR VESICLES RATHER THAN ENVELOPED VIRUSES present in the sample? Equally, HOW CAN VIRAL RESEARCHERS KNOW THAT THEY ARE NOT DETECTING SIMILARLY SIZED NON-VIRAL VESICLES OR EMPTY VECTORS during vaccine production?”
https://www.google.com/amp/s/www.nanoviewbio.com/exosome-blog/2020/5/5/extracellular-vesicles-and-viruses-two-sides-of-the-same-coin%3fformat=amp
“Nowadays, IT IS AN ALMOST IMPOSSIBLE MISSION TO SEPARATE EVs AND VIRUSES by means of canonical vesicle isolation methods, such as differential ultracentrifugation, because they are frequently co-pelleted due to their similar dimension [56,57]. To overcome this problem, different studies have proposed the separation of EVs from virus particles by exploiting their different migration velocity in a density gradient or using the presence of specific markers that distinguish viruses from EVs [56,58,59]. However, to date, A RELIABLE METHOD THAT CAN ACTUALLY GUARANTEE A COMPLETE SEPARATION DOES NOT EXIST.”
Click to access viruses-12-00571.pdf
“The challenge in isolating vesicles is DIFFERENTIATING THEM FROM OTHER TYPES OF MEMBRANE MATERIAL in the cell culture supernatant.”
https://www.news-medical.net/amp/life-sciences/How-are-Exosomes-Isolated.aspx
“But there are many variants in between. WHERE DO VIRUSES START, AND WHERE DO EXTRACELLULAR VESICLES START?”
https://www.quantamagazine.org/cells-talk-in-a-language-that-looks-like-viruses-20180502/
LikeLiked by 1 person
In reply to your comment starting with: ‘You must not understand that the scientific method requires a valid independent…”
Again, cause and effect are two entirely different things. I do not agree with their conclusions whatsoever on cause and effect (i.e. infectiousness). However, I do agree they are able to isolate and characterize the proteins of viruses, enzymes, exosomes, and so forth. When you say “completely separated”, you are acting as if particles the size of viruses can be 100% removed from the fluids by which they are bound. That is a physical impossibility. If you take a rock and isolate it from the dirt that surrounds it, you have a completely isolated rock, and it was likely quite easy to do so because of the size of the rock. Yet, the smaller something gets, the more is required to isolate the object. This is a law of nature and has nothing to do with virology.
With objects as small as viral particles, exosomes, and so forth, damage to those particles becomes a major issue. Complete and total isolation as you are describing is not needed to determine the size, shape, and characteristics of the rock, even if it still has a little dirt around it.
The study you provide concludes:
“Here, we aimed to create awareness that virus preparations may never be pure but rather are contaminated with diverse subpopulations of EVs, and SOME of these EVs may be either indistinguishable from or very similar to so-called defective viruses.”
Further, the study never claims viruses are not real, but affirms it as fact.
It concludes, in part, with the following:
“Understanding of the structure of EVs produced by infected cells, determining their cargo, and deciphering the fine mechanisms by which they affect viral infection are required not only for basic virology but also for translation into therapy.”
Concerning “To study their respective functions…”:
The study states:
“This is very difficult with some viruses, such as retroviruses, because both EVs and retroviruses are comparable in size (EVs ranging from 50 to 100 nm, virions being ∼100 nm) and buoyant density (EVs: 1.13–1.18 g/L; most retroviruses: 1.16–1.18 g/L). Other membrane-derived materials may also have similar characteristics. Therefore, density gradients, which are often used to separate EVs from contaminating protein aggregates on the basis of differences in buoyant densities (40), are not always reliable for separation of EVs from viral particles.”
Thus, proper procedures must be carried out to differentiate. They do not claim such things cannot be done, and even allude to the fact that they can, but that it must be done properly.
LikeLiked by 1 person
“When you say “completely separated”, you are acting as if particles the size of viruses can be 100% removed from the fluids by which they are bound. That is a physical impossibility.”
I agree that the smaller the object gets, the more impossible it is to separate completely yet this is the problem virologists and exosome researchers have locked themselves into trying to claim lab-created nanoparticles have any relevance whatsoever. If they can not separate, they have no valid independent variable. If they can not separate, they can not characterize the particles as they are in a mixture with millions of other particles. They also can not determine which particles are the ones they are supposedly studying if they are in a mixed state. This is just straight up logic you are missing along with an understanding of the scientific method.
“Complete and total isolation as you are describing is not needed to determine the size, shape, and characteristics of the rock, even if it still has a little dirt around it.”
We can see the rock and hold it. We can not do this with “viruses” and exosomes.
“Further, the study never claims viruses are not real, but affirms it as fact.”
I never stated any study I provided claimed “viruses” are not real. In fact, I use their own sources to show the illogical conclusions, contradictions, and the lack of adherence to the scientific method. The fact that they believe in “viruses” does not negate the information I shared from the source.
“Thus, proper procedures must be carried out to differentiate. They do not claim such things cannot be done, and even allude to the fact that they can, but that it must be done properly.”
Where were these procedures carried out properly? Where is the adherence to the scientific method? Please explain how they can characterize the particles without separating them from everything else. How are they able to determine which are the particles they are studying in EM images if there are millions of similar/identical particles within the sample as well?
LikeLike
While we may not be able to touch them or see them directly, at the same time, we cannot see or touch most anything in the body. Surely, you can understand my rock example for what it is. The fact remains, that there must be a mediator between cells to deliver cellular fluids and facilitate communication between cells. Exosomes are merely one such agent that takes this role. At the same time, there must also be a sole solvent factor to break down toxicity when our living microbes are poisoned by modern toxins and can no longer take part in the engulfing of that toxic matter without being poisoned to death. Thus, a solvent construct must absolutely exist to break down substances so cells can rationally handle them.
The body is quite adept at creating solvents and acids. Each cell is a micro-laboratory, with all of the tools to manufacture many types of proteins and variants therein.
Viruses allow cells to dissolve substances without the cell ever making contact with the whole toxin. What is left after viral dissolution is the remnants of a once caustic toxin, which are then more easily managed and removed by cells. It is mainstream science itself that admits that viruses are able to dissolve the walls of cells.
We must take the larger picture into context. None of these entities exist in singular environments in the body. They exist in harmony, with intelligence and cohesion. If we took a human out of the natural world and completely sterilized the body of all bacteria with harsh sterilizers, they would eventually no longer function properly, would become mutated, and die. Likewise, on the macro scale, it is irrational to expect pure isolation and sterilization of an organic particle that cannot be completely removed from its natural environment. To do so would surely mean the destruction of the particle, including misobservation of functions. You can only purify a particle so much before it alters its structure. This is especially true for organic particles.
There are not millions of particles that look like viruses under electron microscopy. Viruses are clearly defined from any other body. You must train yourself to see the differences. When you say they can not determine which particles are which, this is not accurate. For example, you can easily determine an adenovirus particle from influenza virus particles.
Further, if I am not mistaken, I do believe the study you are quoting is referring to vesicles that themselves contain viral particles, not that exosomes and viruses cannot be seen or isolated. As I wrote before, you cannot separate a protein from another protein like an exosome without destroying both. Exosomes may carry viral particles at some point, but viruses and exosomes appear separate much of the time. I already explained how they can conduct characterization.
LikeLiked by 1 person
“The fact remains, that there must be a mediator between cells to deliver cellular fluids and facilitate communication between cells. Exosomes are merely one such agent that takes this role.”
Even if this is true, that does not mean that the particles seen in EM and said to be exosomes are in fact the agents that perform this role. This is nothing but speculation about particles which are picked out as the representative particles in EM. They are most likely nothing but cellular debris. There is no reason to believe otherwise.
“Viruses allow cells to dissolve substances without the cell ever making contact with the whole toxin.”
Where was a “virus” ever properly purified/isolated and proven to function in the way you claim?
“There are not millions of particles that look like viruses under electron microscopy.”
There are supposedly 380 trillion “viruses” in our bodies and 38 trillion bacteria. Who knows how many exosomes are said to be within the body. This is not including other microvesicular bodies, apoptotic bodies, clathrin-coated vesicles, secretory vesicles, glogi and coatomer-coated vesicles, etc. which are all mistaken for “viruses” in EM.
“For example, you can easily determine an adenovirus particle from influenza virus particles.”
Where were either the adenovirus and influenza “viruses” ever purified/isolated in order to determine what they look like?
“I already explained how they can conduct characterization.”
Where? I have yet to see an explanation for how they can characterize anything within a mixed population of particles of the same size, shape, and density.
LikeLiked by 1 person
“Where was a “virus” ever properly purified/isolated and proven to function in the way you claim?”
By your standards, nothing in the body is known. We are not finding any common ground if you cannot even agree with anything I have written. There’s literally nothing I can provide to you if you are already dead-set on believing that none of these things exist. Take a step back for a moment and think about what I am imparting without bias. Some things in life must be theorized and concluded in scientific reasoning and likelihood. You seem to be suggesting that researchers should automatically have all the answers for every facet of every atom of something to conclude it is real. Real science is far more mature than that, even if it is tainted by the modern aura of deception.
If exosomes were called by any other name, you surely would find them faulty as well.
“Even if this is true, that does not mean that the particles seen in EM and said to be exosomes are in fact the agents that perform this role.”
The way one would know that is by observing the components of the vesicle itself and its makeup, like the membrane, shape, size, etc. Simple cellular debris does not exactly appear like vesicles, even though they were originally thought of as cellular debris back in the 80s with little evidence to go on. There are likely other vesicles that researchers have mistaken as debris, which are actually reconstitutions of cellular parts packaged up and used in the creation of new cells. The body recycles when it can.
“Where were either the adenovirus and influenza “viruses” ever purified/isolated in order to determine what they look like?”
By centrifuging a sample, extracting the proper density band containing the virus, and observing it through electron microscopy, then taking photographs.
Your only line of defense thus far has been: “Where was a “virus” ever properly purified/isolated?” There are many photographs and photographs in studies, many of which you yourself have quoted from and referenced, that show viruses. You do not believe viruses exist, so you are already greatly biased. This is not an argument about the supposed infectiousness of these entities but about their actual existence, which is an entirely and vastly different argument. In my view, it distracts from actual errors in virology.
These are the same sort of tactics Kaufman and Cowan (and Lanka) resort to. Their entire modus operandi is to heavily cherry-pick studies to try to prove their conclusions, and they do so in a manipulative manner. It is misrepresenting fundamental science and making it appear as if none of these things exist and that researchers are completely inept at every step. For example, they have set the bar for isolation so high that they can potentially claim nothing exists. Where’s the middle ground?
At the same time, they have absolutely no issue promoting and selling the ‘Analemma water wand’, which promises to ‘restructure’ tap water, and will gladly show you the so-called ‘science’ behind the scam product. In actuality, it will only restructure your wallet. Many of the same people who believe viruses are not real will go to their website and purchase this wand based on their words alone. In hindsight, you cannot turn tap water into water for healthy use in the body, and it is absurd to even make such claims. You do not need a study to prove that fact, but I digress.
My comment: “There are not millions of particles that look like viruses…” was in reference to your previous comment, “How are they able to determine which are the particles they are studying in EM images if there are millions of similar/identical particles within the sample as well?”
My point was that there are not millions of other particles besides viruses that look like viruses. Exosomes and cell debris do not look like viruses—viruses have unique structures, some more so than others.
The most important aspect to understand is, that without viruses, the body would surely die. Our cells need specialized non-living solvents to disassemble/dissolve substances in and around tissues that are not bio-active. Normal cleansers, like bacteria, fungus, parasites, phagocytes, etc, will not survive such non-bio-active toxins for long because they would all be killed. Even on a theoretical basis, you cannot legitimately deny that the body must have such cell constructs. Otherwise, we would all be dead. Think of all of the manmade industrial pollution, waste, and chemicals that the average person is exposed to on a daily basis that is detrimental to all life. Do you suppose the body magically removes all of this using its living agents? How can it? It cannot. It must have specialized, highly specific solvents, period.
LikeLiked by 1 person
“By your standards, nothing in the body is known.”
It’s not my standards. It is the scientific method.
“The way one would know that is by observing the components of the vesicle itself and its makeup, like the membrane, shape, size, etc. Simple cellular debris does not exactly appear like vesicles, even though they were originally thought of as cellular debris back in the 80s with little evidence to go on. There are likely other vesicles that researchers have mistaken as debris, which are actually reconstitutions of cellular parts packaged up and used in the creation of new cells. The body recycles when it can.”
So you are claiming they can figure out what these particles are based on what they look like. In other words, pure speculation.
“By centrifuging a sample, extracting the proper density band containing the virus, and observing it through electron microscopy, then taking photographs.”
The problem, again, is that they admit that there are many particles of the same size, shape, and density that collect together in the band. They can not separate them. Thus, any EM photographs are meaningless as they are picking out whichever particles fits their preconceived idea of what they want their entity to look like.
“There are many photographs and photographs in studies, many of which you yourself have quoted from and referenced, that show viruses.”
Same problem as above. These are images of particles created after heavy alterations in cell culture and TEM preparation. There is no evidence whatsoever that these particles exist in the body as presented nor that they have any function in the body. They are either cellular debris created from the processes the sample is put through or artefacts of TEM imaging. The images are so far removed from reality that they are useless.
“These are the same sort of tactics Kaufman and Cowan (and Lanka) resort to. Their entire modus operandi is to heavily cherry-pick studies to try to prove their conclusions, and they do so in a manipulative manner. It is misrepresenting fundamental science and making it appear as if none of these things exist and that researchers are completely inept at every step.”
You are misrepresenting science by not holding it to the scientific method. Remember these two definitions:
SCIENCE: knowledge or a system of knowledge covering general truths or the operation of general laws especially AS OBTAINED AND TESTED THROUGH SCIENTIFIC METHOD.
https://www.merriam-webster.com/dictionary/science
PSEUDOSCIENCE: Theories, ideas, or explanations that are represented as scientific but that are NOT DERIVED FROM SCIENCE OR THE SCIENTIFIC METHOD.
https://www.oxfordreference.com/view/10.1093/acref/9780199594009.001.0001/acref-9780199594009-e-1007
You are promoting pseudoscience, speculation, and guesswork based on observing random particles and creating functions around them.
“Exosomes and cell debris do not look like viruses—viruses have unique structures, some more so than others.”
First, they are all cell debris. And second, I have repeatedly shown that researchers claim “viruses” and exosomes can not be distinguished:
“PHYSICAL and CHEMICAL CHARACTERISTICS of many EVs, as well as their biogenesis pathways, RESEMBLE THOSE OF RETROVIRUSES. Moreover, EVs generated by virus-infected cells can incorporate viral proteins and fragments of viral RNA, BEING THUS INDISTINGUISHABLE FROM DEFECTIVE (NONINFECTIOUS) RETROVIRUSES.”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4995926/
You seemed to agree in the past:
“It can be said that EXOSOMES ARE INDISTINGUISHABLE FROM VIRUSES in some cases. Exosomes are the same size as viruses (0.1 microns)—and vary in size, but are mostly of similar size.”
https://virusesarenotcontagious.com/exosomes-vs-viruses/
“The most important aspect to understand is, that without viruses, the body would surely die.”
This is a substantial statement. Do you have any evidence supporting this or just speculation?
“Do you suppose the body magically removes all of this using its living agents?”
You are conflating many things and putting them together into a unobservable process. The body does rid itself of waste. However, to claim it is due to “viruses” and exosomes is ridiculous. You have concocted a fantasy using fictional entities that were created in a petri dish by heavy cell altering processes which can only be observed dead.
LikeLiked by 1 person
Jeff,
“The most important aspect to understand is, that without viruses, the body would surely die. Our cells need specialized non-living solvents to disassemble/dissolve substances in and around tissues that are not bio-active. Normal cleansers, like bacteria, fungus, parasites, phagocytes, etc, will not survive such non-bio-active toxins for long because they would all be killed. Even on a theoretical basis, you cannot legitimately deny that the body must have such cell constructs. Otherwise, we would all be dead. Think of all of the manmade industrial pollution, waste, and chemicals that the average person is exposed to on a daily basis that is detrimental to all life. Do you suppose the body magically removes all of this using its living agents? How can it? It cannot. It must have specialized, highly specific solvents, period.”
I’m open to your paradigm in general, but it must accord with Reason in order to be true. The statement you made above may not stand to reason.
The reason that I mistook microsomal monooxygenases as what I thought you were mistaking viruses for being was because they are the major class of enzymes, some of which that have presumably evolved more recently in order to deal with the slew of industrial chemical xenobiotics. And presumably they evolved in humans via HGT via microbes and ultimately via the primordial soup.
Anaerobic microbes themselves have evolved to handle nasty ass shit. To bioremediate it so that aerobic life can repopulate the locale.
I feel that you underestimate the resiliency and redundancies of the living ecology when you say that it absolutely cannot adequately detoxify without what it is that you conceive to be the additional, essential viral enzyme function.
In a previous thread here I explored the body’s biological remediation dynamics.
Here’s a link to my first relevant comment.
https://viroliegy.com/2022/06/05/monkeypox-mania/#comment-3341
If you use the ‘find in page’ function and search for “anaerobe” it will highlight further examinations of the body’s bioremediation abilities. The key thing to understand about the bioremediation of industrial chemicals is that generally they are fat-soluble so they get stored in body fat and must be enzymatically broken down into water soluble toxins so that the anaerobes and the other enzymes can work on them as well as enable the toxins to return to the bloodstream through capillary walls which requires water solubility.
Cheers.
LikeLike
Sorry, Jeff, I cut a sentence short: monooxygenases are the major class of enzymes that deal with fat-soluble toxins.
LikeLike
Going back to your past posts, you clearly claim that exosomes are real. You had even posted some of my writings which you have removed as well. Now, you have done a complete 180 and suddenly changed your stance altogether.
Here, for example, you concluded the following: ““Viruses” are nothing more than exosomes. These are just two competing theories for the exact same particles created from artificial conditions in a lab.” https://viroliegy.com/2021/09/10/viruses-or-exosomes/
I wrote “It can be said that exosomes are indistinguishable from viruses in some cases.” nearly three years ago—”…in some cases.”
I also wrote this on June 29th, 2022: “It may correctly be stated that viruses appear similar to other agents from a distance, as all other agents in the body do. Therefore, taking adequate time to identify is needed.” https://jeffgreenhealth.substack.com/p/are-viruses-merely-cell-debris.
And, in my first comment to you, I wrote “On their surface, many cells and entities in the body appear to be similar or the same, until closely observed.”
These statements are still true. You are merely taking one statement I made out of its context. You are far too defensive to have a legitimate conversation with and you addressed nothing of import from my previous reply other than to deflect and deny.
“These are images of particles created after heavy alterations in cell culture and TEM preparation. There is no evidence whatsoever that these particles exist in the body as presented nor that they have any function in the body.”
Then you are basically admitting cell alteration elicits cells to produce viral proteins. If cells can produce such proteins in cell culture, then they can do so in the body. I have concluded that your entire stance is to chase your own tail. It is as if you are fighting with yourself. I have nothing to fight with you over, but I simply disagree with your broad and generalized denial of all things science.
With regard to your comments on isolation, you are wholly mistaken. You expect those conducting isolation to separate viruses 100% from their fluids to be able to observe them, which is impossible. You do not use the term “100%”, but that is obviously what you allude to. You are using that minor negative to try to prove your denialism. You are claiming that viruses are merely mistaken as cell debris, even though viruses have apparent structures and cohesion in their form when closely observed. Of course, you do not believe any of this exists anyway.
“I have repeatedly shown that researchers claim “viruses” and exosomes can not be distinguished”
You have done no such thing. The very study in your post here states the following: “…and some of these EVs may be either indistinguishable from or very similar to so-called defective viruses.” p.9159
“Some of”, and in “defective viruses”, which are incomplete viral structures. Further, retroviruses are only one type of virus. Nowhere does it state exosomes cannot be distinguished from all other complete viruses or vice versa, nor does it state that this occurs in every situation by every researcher.
Finally, you baselessly claim I am “promoting pseudoscience”, yet you are on here claiming antibodies, exosomes, viruses, and everything else attached to them are all fake and non-existent, with literally no substitute to take their place. Your argument has no basis in reality. In fact, your stance is that nothing can be proven or known. Essentially, you can deny viruses exist, and by doing so you do not have to address the complexities of so-called infectiousness, symptoms, isolation, or anything else surrounding them for that matter. You have wrapped it all into a nice neat box with a bow on top.
LikeLiked by 1 person
“Going back to your past posts, you clearly claim that exosomes are real. You had even posted some of my writings which you have removed as well. Now, you have done a complete 180 and suddenly changed your stance altogether.”
Nowhere did I state that exosomes exist. In fact, in the very post you shared, I stated this:
“Viruses” are nothing more than exosomes. These are just TWO COMPETING THEORIES FOR THE EXACT SAME PARTICLES CREATED FROM ARTIFICIAL CONDITIONS IN A LAB. One views them as invaders from outside the body while the other envisions them as beneficial messengers from within. The truth is that these particles are not pathogenic, originate from material within the human body, and have had VARIOUS UNPROVEN THEORETICAL FUNCTIONS APPLIED TO THEM. It is even a fair question to ask WHETHER OR NOT THESE PARTICLES ACTUALLY EXIST WITHIN THE BODY IN THE STATE THEY ARE PRESENTED IN RATHER THAN JUST BEING BY-PRODUCT FROM THE CELL DESTROYING METHODS USED TO CREATE THEM. The only thing for certain is that ‘viruses” and exosomes are the exact same particles.”
https://viroliegy.com/2021/09/10/viruses-or-exosomes/
“And, in my first comment to you, I wrote “On their surface, many cells and entities in the body appear to be similar or the same, until closely observed.”
You are basing your views on being able to tell these particles apart based on only appearance. If the particles in question are not completely isolated from all other similar/identical particles that are said not to be them, there is no way that the researcher can know that they picked the right particles as the culprit even upon close observation. They are just pointing and declaring at whatever ones suit their needs.
“Then you are basically admitting cell alteration elicits cells to produce viral proteins. If cells can produce such proteins in cell culture, then they can do so in the body.”
I admit that the cell culture process destroys the cell and breaks it apart. I also state that the purification methods break the particles into smaller and smaller pieces. The EM preparations further alter the sample. There is no way to claim that the particles observed in EM have any relation to what may have been originally in the sample. It is so far removed from reality that it is meaningless to make claims and assign functions onto these heavily altered dead particles and/or potential artefacts.
“I have nothing to fight with you over, but I simply disagree with your broad and generalized denial of all things science.”
This is because you do not seem to understand the scientific method. It requires the observation of a natural phenomena as well as a valid independent variable that can be manipulated and varied. It also requires valid and proper controls to be carried out. Virology does not follow the scientific method and neither does exosome research.
“You are claiming that viruses are merely mistaken as cell debris, even though viruses have apparent structures and cohesion in their form when closely observed. Of course, you do not believe any of this exists anyway.”
What are “virus” particles and how were they determined to be “virus” particles in a mixed culture with similar/identical particles? Go back to any foundational paper for any “virus.” None of the studies purify nor isolate any particles assumed to be “virus” nor are the particles ever proven pathogenic. They are creations stemming from tissue/cell cultures. Again, the researchers point and declare at whatever particles they want to represent their fictional entity.
“Nowhere does it state exosomes cannot be distinguished from all other complete viruses or vice versa, nor does it state that this occurs in every situation by every researcher.”
Many sources claim this:
“Unless more specifically defined, it is currently VIRTUALLY IMPOSSIBLE TO SPECIFICALLY SEPARATE AND IDENTIFY EVs that carry viral proteins, host proteins, and viral genomic elements FROM ENVELOPED VIRAL PARTICLES that carry the same molecules.”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4995926/
“THE GENERIC CHARACTERIZATION OF EXTRACELLULAR VESICLES COULD ALSO BE USED AS A DESCRIPTOR OF ENVELOPED VIRUSES, highlighting the fact that extracellular vesicles and enveloped viruses are similar in both composition and function. THEIR HIGH DEGREE OF SIMILARITY MAKES DIFFERENTIATING BETWEEN VESICLES AND ENVELOPED VIRUSES IN BIOLOGICAL SPECIMENS PARTICULARLY DIFFICULT.”
“However, as previously reviewed, exosomes and viruses do not conform to strict definitions [22]. Intermediate particles exist on the spectrum between virus and exosome that contain both host and viral components, MAKING IT NEARLY IMPOSSIBLE TO CLASSIFY THESE VESICLES AS EITHER DEFECTIVE VIRUSES OR EXOSOMES THAT CONTAIN VIRAL COMPONENTS [22]. INTERMEDIATE PARTICLES ARE OFTEN CLASSIFIED AS A VIRUS OR EXOSOME DERIVATIVE, DEPENDING ON THE PREFERENCE OF THE INVESTIGATOR, but once these vesicles deviate from strict definitions they may be more accurately defined as an assortment of lipid-encased particles THAT CANNOT BE EASILY DIFFERENTIATED [22].”
https://www.microbiologyresearch.org/content/journal/jgv/10.1099/jgv.0.001193?crawler=true
“This makes EV CONCEPTIONALLY AND BIOCHEMICALLY SIMILAR TO VIRUSES [1,18] (Figure 1C). The formation and egress of microvesicles and exosomes share similarities to virus biogenesis, such as HIV [24] and enveloped hepatitis A viruses (HAV) [25] (Figure 1), respectively. In fact, VIRUSES HAVE BEEN THOUGHT OF AS EMERGING FROM EXOSOMES OR VICE VERSA [26]. ANY EV CAN PLAY AN ANALOGOUS ROLE TO A VIRUS PARTICLE in the functional transfer materials from one cell to the next, regardless of the class.”
https://www.google.com/url?sa=t&source=web&rct=j&url=https://www.mdpi.com/1999-4915/12/9/917/pdf&ved=2ahUKEwi8x6SNvcnxAhWGW80KHfVPB3EQFjAMegQIERAC&usg=AOvVaw3kccThKbbHOPZasZ_5KBWb
“Essentially both are encapsulated by a lipid membrane, rich in membrane proteins and contain nucleic acids. The composition of the lipid membrane is very similar, with some viruses (e.g. HIV and HepC) shown to have tetraspanins on their surface. Viruses can vary greatly in size but there is certainly a significant number in the sub 200 nm range, SARS-CoV2 is itself around 60-140 nm in diameter. There is some evidence that extracellular vesicles can also have a protein corona which may contribute towards functional effects. THIS, ALONG WITH SIZING AND DENSITY SIMILARITIES MAKES IT HARD TO DISTINGUISH BETWEEN THE TWO USING EM.
HOW CAN WE BE SURE that we are isolating and quantifying extracellular vesicles rather than enveloped viruses present in the sample? Equally, HOW CAN VIRAL RESEARCHERS KNOW that they are not detecting similarly sized non-viral vesicles or empty vectors during vaccine production?”
https://www.google.com/amp/s/www.nanoviewbio.com/exosome-blog/2020/5/5/extracellular-vesicles-and-viruses-two-sides-of-the-same-coin%3fformat=amp
“Since VESICLES RESEMBLE VIRUSES, the question of course is whether the first extracellular vesicles were primitive viruses and the viruses learned from extracellular vesicles or vice versa.”
“Viruses can replicate and vesicles cannot. But there are many variants in between. WHERE DO VIRUSES START, AND WHERE DO EXTRACELLULAR VESICLES START?”
Leonid Margolis
https://www.quantamagazine.org/cells-talk-in-a-language-that-looks-like-viruses-20180502/
“Finally, you baselessly claim I am “promoting pseudoscience”, yet you are on here claiming antibodies, exosomes, viruses, and everything else attached to them are all fake and non-existent, with literally no substitute to take their place.”
Yes, you are pepetuating pseudoscience as you are basing your theory on evidence obtained without adherance to the scientific method. I have looked at the foundational evidence for antibodies, exosomes, and “viruses” and have shown that these studies do not adhere to the scientific method. Thus, it is easy to say that there is no scientific basis for the existence of these fictional entities. All the researchers do is create particles and declare whichever one fits their agenda as the culprit and assign a name and fuction to it. You are doing the same.
LikeLike
You are still not properly differentiating between cause and effect and isolation, which again, are each different processes. You are combining them into one process. Importantly, you do not need to show the supposed cytopathic effects of a particle in order to isolate it. That is secondary to isolation and comes afterward.
With regard to exosomes and viruses being nearly impossible to isolate from each other—you are misreading the study from which you are quoting. It clearly reads that viral particles that exist inside exosomes cannot be separated from one another. Researchers do not need to do this to observe viruses anyway because both particles appear in separation in samples. This is done in order to determine the cargo of an exosome, which is part of the topic of the study.
Furthermore, your statements about viruses being merely cellular debris, as you continually rehash, is missing one very important observation, among many. That is, if viruses were simply cellular debris, pictures of viruses would show them without cohesive form; without apparent structure. You see an equilateral distribution of surface glycoproteins on viruses—something only an intelligent formation by a living organism could manufacture—not the more simple structures and greater various sizes of cellular debris. You may then claim the pictures themselves are fake, but that would be absurd since they are clearly real photographs of organic particles.
Vesicles are merely packages of material that are composed inside the cytoplasm of cells, which are either recycled for eventual use by the cell or sent off as extracellular vesicles wherein they gain a lipid membrane by nature of the exocytosis of the particle from the cell wall when they bud off. Some viruses, such as enveloped viruses, also gain their membrane by the same pathway.
Viruses begin in the nucleus of cells. Exosomes do not. They contain entirely different inner core makeups.
Viruses are densely packed with tightly coiled mRNA, which is obtained by the cell nucleus during DNA to RNA transcription using chromosomes. The final viral formation is completed in the cytoplasm by ribosomes, wherein translation of RNA to mRNA takes place. If viruses leave the cytoplasm as unfinished vesicles, it is to transmit the necessary components for less than fully vital cells to manufacture proteins. Thus, incomplete “defective” viruses can become vesicle-like. Such so-called viruses are mere vesicles and not complete viral structures.
All of this is able to be determined by observing formations after they leave the nucleus and/or cell. Chromosomes, for example, cannot be seen in the nucleus under microscopy. However, when cells divide, their appearance is readily visible because their DNA becomes so tightly packed. You may pull up videos of the process. The same is true for the formation of other proteins, like viruses. Simply suggesting that researchers are completely inept in every way is a great disservice to the men and women who spend their lives studying the great complexities of the body and does not seek to correct any errors in medical science. You are making overly broad generalizations that are circular in logic. As I stated before, existence and functionality are entirely different, and by combining these two you will never find the truth of the matter.
To disprove your ideation on vesicles: A cell undergoing apoptosis will release all of its inner parts, and many of them, once they leave the cell, will by the very nature of cellular decay form vesicles—called blebbing, which causes apoptotic membrane protrusions that form into vesicle bodies—that are eventually reestablished in other cells, if they can be utilized and are still bio-active. This is essentially what exosomes are. You are suggesting these vesicles are merely cellular debris with no use, or worse, that they simply do not exist. True cellular debris appears as a fine particulate, not as defined vesicles.
I have video of this near the end of my presentation ‘Viral Misconceptions’ where you can clearly see this breakdown particulate, along with vesicle secretion, from a dying neutrophil.
If cells did not recycle their contents, the body would be incredibly inefficient, and the body is incredibly efficient.
LikeLiked by 1 person
“Debris” in this context just means they have no basis for slapping the label they slapped on an emission of a dying cell based on a story, such as a story about its shape (e.g., spiky ball). “Debris” doesn’t mean it has no purpose or won’t be taken up by other cells, etc. It means the observations do not warrant any special label being placed on the object. It’s a reference to the “point and declare” method of virology.
There’s no need for virologists to be bumbling fools to have virology be a pseudoscience. All that they have to be is unwilling to question the fundamentals of the field, such as those of Nobel laureate Enders. Indeed when pressed on the scientific basis for their methods, the usual response is, for example, “This is how virus isolation is done in virology.” They do not talk about isolating variables and so on. They simply appeal to common practice.
Most don’t even seem to question the dogma that has built up since Enders; they simply take it as given and proceed from there. Any intelligence they have doesn’t matter if they aren’t applying it to the whole process. As we saw with the first “Covid-19” papers, often a given researcher will intelligently and conservatively give a conjecture, a “maybe,” that is then referenced in subsequent papers by different researchers as an established truth.
There is never a guarantee nor even an expectation that humans in groups will act intelligently, even if they are each intelligent themselves. The only exceptions are markets, and in part due to huge government subsidies most parts of science are far removed from market tests. The test is what gets you promoted, tenure, grants, awards, invitations, etc.
This is why institutional science in general is deeply corrupt, with politicized fields like physiology being especially so. Groupthink and “go along to get along” is absolutely the norm even in fields as dispassionate and apolitical as mathematics. When even questioning the fundamentals of set theory will render you persona no grata as far as promotion and tenure and awards, the state microbiology and physiology are in shouldn’t be surprising.
LikeLiked by 1 person
“You are still not properly differentiating between cause and effect and isolation, which again, are each different processes.”
I’m not sure why you are having a hard time understanding this. Isolation must come first in order to obtain the independent variable (i.e. the particles assumed to be exosomes/”viruses” free of all other kinds of particles) which is then used to determine cause and effect. You must also have purified/isolated particles free of everything else in order to properly characterize the particles you are supposed to be studying. If you do not have the particles by themselves, then there is no way to determine specific proteins, antibodies, etc. as they could belong to any of the other substances/contaminants in the sample.
“It clearly reads that viral particles that exist inside exosomes cannot be separated from one another. Researchers do not need to do this to observe viruses anyway because both particles appear in separation in samples.”
It clearly is stating that the particles claimed to be exosomes and “viruses” can not be separated/differentiated due to their similar size, shape, and density. Both particles do not appear in separation. I have shown numerous statements from researchers claiming it is impossible to separate them. One more time:
“Nowadays, it is an almost IMPOSSIBLE MISSION TO SEPARATE EVs AND VIRUSES BY MEANS OF CANONICAL VESICLE ISOLATION METHODS, such as differential ultracentrifugation, because they are frequently co-pelleted due to their similar dimension [56,57]. To overcome this problem, different studies have proposed the SEPARATION OF EVs FROM VIRUS PARTICLES by exploiting their different migration velocity in a density gradient or using the presence of specific markers that distinguish viruses from EVs [56,58,59]. HOWEVER, TO DATE, A RELIABLE METHOD THAT CAN ACTUALLY GUARANTEE A COMPLETE SEPARATION DOES NOT EXIST.”
Click to access viruses-12-00571.pdf
“That is, if viruses were simply cellular debris, pictures of viruses would show them without cohesive form; without apparent structure.”
Pictures of “viruses.” Why do you think pictures of heavily altered random dead particles is valid evidence? Please explain how they obtained the image of the first ever human “virus” if it was in a mixed sample full of other “viruses,” exosomes, apoptotic bodies, and other multivesicular vesicles. Without begging the question, please share how were they able to determine the particles they chose were the “virus” they were studying versus one of the many similar/identical particles in the sample.
“Viruses begin in the nucleus of cells. Exosomes do not. They contain entirely different inner core makeups.”
How can they determine the core of either “virus” or exosome if they can not separate the particles from each other and study either independently?
“All of this is able to be determined by observing formations after they leave the nucleus and/or cell.”
Observing formations tells nothing. You are fixated on static EM images that have been used to create a fictional story of how these particles form and function. Looking at images of heavily altered samples of dead particles tells nothing about whether or not these particles are in the human body in the form they are imaged in nor if or how they may function within a living body. It is nothing but creative writing.
LikeLike
You go sentence by sentence instead of understanding the whole of what I write. So, I can also apply that to you as well.
“Isolation must come first in order to obtain the independent variable (i.e. the particles assumed to be exosomes/”viruses” free of all other kinds of particles) which is then used to determine cause and effect.”
I’m not interested in cause and effect. I am interested in your claims about isolation not taking place. You did not address my point, which is, that you are combining both isolation and CPE together, and you cannot do that. They are separate processes.
“You must also have purified/isolated particles free of everything else in order to properly characterize the particles you are supposed to be studying.”
You do not. Your definition of “free of everything else” is 100% isolation, which is not possible.
In a previous thread, you wrote:
100% isolation is not needed to characterize particles or nothing in the body would be characterized. How can you not concede this fact? You write about others having “intellectual honesty”. Well, where is yours?
“If you do not have the particles by themselves, then there is no way to determine specific proteins, antibodies, etc. as they could belong to any of the other substances/contaminants in the sample.”
Particles are quite pure after proper centrifugation as described in isolation technical manuals:
Lawrence, Janice & Steward, Grieg. (2010). Purification of viruses by centrifugation. 10.4319/mave.2010.978-0-9845591-0-7.166.
“It clearly is stating that the particles claimed to be exosomes and “viruses” can not be separated/differentiated due to their similar size, shape, and density. Both particles do not appear in separation.”
Yes, they do. Viruses and vesicles appear as their own individual bodies. The study you quote from writes about them being joined, which I already clearly address in my previous comments. It is “almost impossible” to separate combined exosomes and viruses.
See here:
Can you not read that for what it states? “EVs that carry viral proteins…” Viruses do not always appear in exosomes—they appear on their own, I.e. single.
“How can they determine the core of either “virus” or exosome if they can not separate the particles from each other and study either independently?”
Refer to my point above.
“Observing formations tells nothing.”
It tells us a lot more than nothing. It tells us structure, through which one learns to characterize objects. How can you be so boldened to claim that “Observing formation tells us nothing.”
I’m not actually fixated on anything. You are, and you are fixed on your theory of non-existence no matter the evidence presented. I am well content in my rebuttals to you. I am not being combative in any way, as you are, because I am content with the correctness of my information, and it is something I have labored over for a period of many years. Your exosome theory is merely a couple of weeks old.
You literally discarded a wealth of information I imparted in my previous post as if it was nothing, which is why I will likely refrain from taking the time to converse with you from here on since you cannot converse in good faith.
LikeLiked by 1 person
The appeal to “I thought a lot about my theory” falls flat when it becomes obvious you haven’t read through Mike’s material much. For example, this article from over 9 months ago makes the same case, that there’s no evidence that “exosomes” and “viruses” aren’t just labels slapped on shapes of similar cellular excretions or breakdown products and that purification to the level necessary for the characterization that would enable virology to get off the ground has never been achieved.
https://viroliegy.com/2021/09/21/the-purification-problem/
As to your quote, merely because a method is commonly used in virology “to purify viral particles” doesn’t mean it succeeds. As that article and several others here amply demonstrate.
You accuse Mike of selective reading and not seeing the big picture but that seems like projection as that’s exactly what you’re doing. You are given quotes many of which definitively say “exosomes” and “viruses” cannot be separated with confidence, and you choose to comment only only those quotes that give you some wiggle room while ignoring the rest. And you ignore the big picture in failing to notice how hilariously convenient of a rescue device it is that one of these objects is said to sometimes contain the other.
When you’ve studied enough scientific bullshitting in enough fields a pattern emerges and virology happens to fit it perfectly, right along with the very idea of “exosomes” and containment.
LikeLiked by 1 person
“I’m not interested in cause and effect.”
Then it is clear you do not care about science and that you are concerned only with pseudoscience:
“Scientists seek to understand the relationships and intricacies BETWEEN CAUSE AND EFFECT in order to predict outcomes of future or similar events. To answer these questions and to help predict future happenings, SCIENTISTS USE THE SCIENTIFIC METHOD – a series of steps that lead to answers that accurately describe the things we observe, or at least improve our understanding of them.”
https://extension.unr.edu/publication.aspx?PubID=4239
Science: knowledge or a system of knowledge covering general truths or the operation of general laws especially as OBTAINED AND TESTED THROUGH SCIENTIFIC METHOD.
https://www.merriam-webster.com/dictionary/science
Pseudoscience: Theories, ideas, or explanations that are represented as scientific but that are NOT DERIVED FROM SCIENCE OR THE SCIENTIFIC METHOD.
https://www.oxfordreference.com/view/10.1093/acref/9780199594009.001.0001/acref-9780199594009-e-1007
“You did not address my point, which is, that you are combining both isolation and CPE together, and you cannot do that.”
I did address this but it seems you are unable to understand. CPE is the way virologists determine if a “virus” is present in a sample. However, it can be caused by numerous factors such as bacteria, Mycoplasma, parasites, amoeba, antibiotics, antifungals, passaging, environmental conditions, cell age, etc. If the particles assumed to be “viruses” are not separated from everything else and the variables are not controlled, then other factors/variables can be used to explain CPE other than a “virus.” In order to determine it was the “virus” particles, and only those particles, causing the CPE, complete isolation (free of everything else and all other factors/variables) must be obtained.
It is a moot point anyways as you are continuing to beg the question. Please, share how any original particle associated with any “virus” was determined to be the representative particle if it was in a mixed state with millions of similar/identical particles.
“100% isolation is not needed to characterize particles or nothing in the body would be characterized.”
We are dealing with nanoparticles estimated to be in our bodies in the trillions. If you can not individually separate them, all you are doing is pointing and declaring at EM images the particle you want as your representative and assigning functions to it.
“Particles are quite pure after proper centrifugation as described in isolation technical manuals:”
What is “quite pure?” What is the allowable level of impurity? How many other particles of the same size, shape, density are allowed? If other particles are allowed, how are the ones being studied differentiated and distinguished if all particles are similar and/or identical?
There is either pure or impure. Isolated or not isolated. You are playing the same definition game vitologists do:
“In these terms, PURITY MEANS THE DEGREE OF FREEDOM FROM NONVIRAL COMPONENTS, or, conversely, the extent to which viral particles show gross physicochemical homogeneity. NO SINGLE TEST IS SUFFICIENT TO ESTABLISH THIS TYPE OF PURITY,”
“It is obvious, of course, that IMPURITIES WILL ESCAPE DETECTION IF THEY HAVE THE SAME SIZE AND SHAPE AS THE VIRUS PARTICLES, or if they are below the size resolved by the microscope. Also, particles present in small number but large in mass are easily overlooked, owing to sampling difficulties (Lauffer 1951).”
“In summary, NO SINGLE CRITERION OF PURITY IS SUFFICIENT TO ESTABLISH THE HOMOGENEITY OF A PREPARATION OF VIRUS.”
https://www.google.com/url?sa=t&source=web&rct=j&url=https://link.springer.com/content/pdf/10.1007/978-3-642-85899-4_2.pdf&ved=2ahUKEwi04tPs_7DvAhWOZs0KHZp1BC0QFjANegQIFxAC&usg=AOvVaw0Fx51ebi9lAsov0RyK1wyl
“Viruses and vesicles appear as their own individual bodies.”
I’m only going to post this one more time. I have provided numerous examples where they are clearly talking about not being able to separate/isolate “viral” particles from those claimed as exosomes, not the particles being composed of similar elements:
“Nowadays, it is an almost IMPOSSIBLE MISSION TO SEPARATE EVs AND VIRUSES BY MEANS OF CANONICAL VESICLE ISOLATION METHODS, such as differential ultracentrifugation, because they are frequently co-pelleted DUE TO THEIR SIMILAR DIMENSION [56,57]. To overcome this problem, different studies have proposed the SEPARATION OF EVs FROM VIRUS PARTICLES by exploiting their different migration velocity in a density gradient or using the presence of specific markers that distinguish viruses from EVs [56,58,59]. HOWEVER, TO DATE, A RELIABLE METHOD THAT CAN ACTUALLY GUARANTEE A COMPLETE SEPARATION DOES NOT EXIST.”
Click to access viruses-12-00571.pdf
“Can you not read that for what it states? “EVs that carry viral proteins…” Viruses do not always appear in exosomes—they appear on their own, I.e. single”
They are discussing EVs with “viral” proteins. However, there are no “viral” proteins. These are the same particles. They are not discussing separating “viral” components from EVs, they are discussing how closely related they are which makes it difficult to separate them as distinct entities. From the conclusion:
“A growing body of evidence indicates that cells infected with enveloped or nonenveloped viruses release EVs that contain viral components. Here, we aimed to create awareness that virus preparations may never be pure but rather are contaminated with diverse subpopulations of EVs, and some of THESE EVs MAY BE EITHER INDISTINGUISHABLE FROM OR VERY SIMILAR TO SO-CALLED DEFECTIVE VIRUSES.”
In other words, these “exosomes” are the exact same particles called “viruses” but they are claimed to be defective. The “viral” samples will always be impure as exosomes, which are indistinguishable, are present. This is called a rescue device.
“It tells us a lot more than nothing. It tells us structure, through which one learns to characterize objects.”
These are interpretations of heavily altered samples of dead particles. Familiarize yourself with the work of Harold Hillman and you will understand why looking at EM tells you nothing about what is inside a living organism:
“For example, most cytologists know, but readers of elementary textbooks do not, that when one looks at an illustration of an electron micrograph: an animal has been killed; it cools down; its tissue is excised; the tissue is fixed (killed); it is stained with a heavy metal salt; it is dehydrated with increasing concentrations of alcohol; it shrinks; the alcohol is extracted with a fat solvent, propylene oxide; the latter is replaced by an epoxy resin; it hardens in a few days; sections one tenth of a millimetre thick, or less, are cut; they are placed in the electron microscope, nearly all the air of which is pumped out; a beam of electrons at 10,000 volts to 3,000,000 volts is directed at it; some electrons strike a phosphorescent screen; the electron microscopists select the field and the magnification which show the features they wish to demonstrate; the image may be enhanced; photographs are taken; some are selected as evidence. ONE CAN IMMEDIATELY SEE HOW FAR THE TISSUE HAS TRAVELLED FROM LIFE TO AN ILLUSTRATION IN A BOOK.”
https://www.big-lies.org/harold-hillman-biology/what-price-intellectual-honesty.htm
“During a research career lasting more than 50 years, I HAVE CONCLUDED THAT THE FOLLOWING PROCEDURES ARE UNSUITABLE FOR STUDYING THE BIOLOGY OF LIVING CELLS IN INTACT ANIMALS AND PLANTS: subcellular fractionation; histology; histochemistry; electron microscopy; binding studies; use of ligands; immunocytochemistry; tissue slices; disruptive techniques; dehydration; deep freezing; freeze drying; boiling; use of extracellular markers; receptor studies; patch clamp measurements; inadequate calibrations”
“The nuclei, containing the nucleoli, and the mitochondria, are the only structures seen by light microscopy in the cytoplasm of living cells.
All the other structures claimed to be present are only seen in electron micrographs of dead tissue, or in fixed stained histological sections.
IF THEY EXISTED, the cytoplasm would be almost solid with endoplasmic reticulum, cytoskeleton, Golgi bodies, lysosomes, peroxisomes, contractile proteins and stress fibres.”
https://library-of-atlantis.com/harold-hillman/
“I am not being combative in any way, as you are, because I am content with the correctness of my information, and it is something I have labored over for a period of many years.”
I’m not sure how asking you for validation is combative. You argument assumes these things called “viruses” and exosomes exist and that the functions have been determined. You are begging the question. It is clear you have not looked into the foundational evidence. It is also clear that you are unfamiliar with the scientific method and that you believe EM images are valid proof, irregardless of how they were obtained.
Unlike you, I do not have a “theory.” I am just stating facts. The EM images you are so beholden to are nothing but static images of heavily altered samples in a dead state. To claim function as well as any biological role inside a living organism for cellular debris/artefacts is very unscientific.
LikeLike
Clarifire
“The appeal to “I thought a lot about my theory” falls flat when it becomes obvious you haven’t read through Mike’s material much. For example, this article from over 9 months ago makes the same case, that there’s no evidence that “exosomes” and “viruses” aren’t just labels slapped on shapes of similar cellular excretions or breakdown products and that purification to the level necessary for the characterization that would enable virology to get off the ground has never been achieved.
https://viroliegy.com/2021/09/21/the-purification-problem/”
The fundamental reason — under natural law — that virology never got off the ground is that intelligent patterning by metaconsciousness (MC in my ‘home cosmology’), of natural law, does not admit submicroscopic terrorists without boxcutters to the party. That’s the first inch they can’t have.
Mike’s article, here, and the argument you’re making on his behalf, is merely the standalone deconstruction of virologists’ partial efforts to establish virology by looking at the light-based aspect of the holograms of these ‘outside bodies’ with different kinds of optical magnifying glasses, which is but one of the three main tools they use to traingulate the existence and function of these structural outside-bodies.
The second toolset they use is reason-based chemical eyes, to look at the combined electromagnetic and thermodynamic ‘biochemical’ aspect of the holograms of the outside-bodies. This ‘biochemical’ aspect is how true, industrial Reason has come to object-model the dynamic, reactive Sky as animism calls it, which is the Energy-field symbiont that the consciousness-field symbiont (Spirit) animates into form and AS UNSEEN to the visible spectrum available to the human eye. Biochemistry is Reason modeling the dynamic, roiling nature of Sky and Spirit in symbiosis. When they purify outside-bodies to a high degree they then analyze the chemicals and by FARMING these intelligent life-bearing ‘acids’ (hyperdynamic, intelligent, primordial Sky/Energy Beings) that love nothing more than to make meaningful connections, these farmers of the origins of biological life record the populations of nucleic-acid compounds and non-nucleic acid compounds, among other recordings that they make, and they begin to triangulate form and function. It’s not rocket science, it’s just made possible by a lot of money, machinery, and human MC focus. And flat-out denying this is one of the prices to pay for a postmodernism that knows not how to build in truth because the truth has been shattered, so the deconstructionist ironically hangs his hat on the same hammer that methodically shattered the truth over the centuries because the deconstructionist only wants to shatter that which shattered the truth – because the truth is done shattered already. The ‘truth’ becomes more shattering instead of the building/evolving process which was always the holistic truth of mature ecologies.
Mike’s hard denial of the function of nucleic acids, and your active doubt about them, Clarifire, one way or the other puts the growth of this blog on hold, IMO. Biochemistry needs to be, once and for all, truthfully debunked or recognized, one way or the other. Otherwise the blog cannot hold a legitimate position on the holistic validity of outside-bodies containing nucleic acids which are the chemicals that industrial Reason has assigned as being responsible for biological genesis; the blog cannot say that these bodies have no genesis function if the blog cannot successfully deconstruct the claimed function of the chemical eyes toolset, which requires understanding the toolset for what it is and how it functions symbiotically with Reason’s patterning/triangulating function.
LikeLiked by 1 person
Reante, it seems we agree that nucleic acids would be a nice direction for the blog to explore, if Mike is interested. Whether PCR can farm in the way you describe I am skeptical but I’m not as knowledgeable about PCR as I should be so I will hold a place in my mind for the possibility.
LikeLiked by 1 person
Yes, I would definitely like to break down the foundational evidence for nucleic acids in the future. It is on my ever-expanding list. 😉
LikeLike
Now, to address some of your statements and correct your errors.
You stated:
“There is either pure or impure. Isolated or not isolated. You are playing the same definition game vitologists do”
Your definition of ‘pure’ simply does not exist anywhere in nature. Pure and impure, as you use the terms, signify your lack of understanding concerning the nature of isolation itself and the rules of the natural world. I have already explained these errors in some of my first replies to you which you continue to ignore—very pure samples may be obtained through multiple passes of filtration. Your statements suggesting viruses must be 100% pure to be isolated is a misnomer. 100% isolation of any object does not exist, especially organic tissues.
Furthermore, when you state we have trillions of viruses in the body, we do, but they are largely bacteriophages, which are very large in comparison to influenza virus, coronavirus, and so forth. These are easily distinguished by researchers due to their structure. You left this out of your comments.
Concerning this statement you made:
“They are not discussing separating “viral” components from EVs, they are discussing how closely related they are which makes it difficult to separate them as distinct entities.”
This is incorrect. In both of the studies in question, it states that defective viruses may appear similar to exosomes because viruses in the defective state are incomplete viruses with vesicle-like structures. You also falsely claim that the studies do not write of viral proteins being inside exosomes.
In reference to review ‘The Role of Extracellular Vesicles as Allies of HIV, HCV and SARS Viruses’
This establishes the scope of the review. Enveloped viruses appear similar to exosomes as well because they contain an outer membrane which they gain from budding from the cell wall.
Therefore, the review is pertaining to enveloped viruses, as well as defective viruses—not non-enveloped viruses or any other. As well, it pertains to exosomes containing and transporting viral parts inside them. In all cases, their usage of the word “indistinguishable” applies only to these areas. You are completely misreading these studies.
In reference to study ‘Extracellular vesicles and viruses: Are they close relatives?’
And
This is because EVs will capture material in a cell, including viruses and/or their parts, and transport them out of the cell. This does not in any way suggest these two particles are the same particle, and neither studies suggest this whatsoever. The “relationship” in this case between exosomes and viruses deals with the similarities between enveloped viruses and defective vesicle-like incomplete ‘viruses’—no other virus. Anything else and you are adding to what these studies write. If what you claim is true, both studies would clearly conclude that all viruses appear as exosomes. As I have just proven, what you suggest is not the basis of either study.
And finally, to settle the matter, the same study states:
“noninfectious”, which means defective viruses—not all viruses. As stated prior, noninfectious viruses (defective) and ‘infectious’ viruses (non-defective) appear completely different in form and are more easily recognized from one another.
LikeLiked by 1 person
“Your definition of ‘pure’ simply does not exist anywhere in nature.”
It is not my definition of pure. It is THE definition of pure:
“not mixed or adulterated with any other substance or material.
without any extraneous and unnecessary elements.
free of any contamination.”
https://www.google.com/search?q=pure+definition&sxsrf=ALiCzsYCs2v554loHxIzJYB1j7xZi08TqA%3A1657150803395&ei=Ux3GYtbfF6_JptQPycq7mAk&oq=&gs_lcp=ChNtb2JpbGUtZ3dzLXdpei1zZXJwEAEYBDIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzINCC4QxwEQrwEQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJzIHCCMQ6gIQJ0oECEEYAFAAWABgqRVoAnABeACAAQCIAQCSAQCYAQCgAQGwAQ_AAQE&sclient=mobile-gws-wiz-serp
“Furthermore, when you state we have trillions of viruses in the body, we do, but they are largely bacteriophages, which are very large in comparison to influenza virus, coronavirus, and so forth.”
Please share verification of your claim. The 380 trillion “viruses” said to be within us are supposedly unknown.
“Most viruses REMAIN UNDISCOVERED, REFERRED TO BY SOME SCIENTISTS AS ‘VIRAL DARK MATTER ’. Despite this, they occur through all parts of our body. A study led by Dr Kei Sato from the University of Tokyo published in June 2020 found viruses in human tissues including the brain, blood, kidney and liver.”
https://www.google.com/amp/s/www.sciencefocus.com/the-human-body/the-human-virome-the-trillions-of-viruses-inside-your-body-keeping-you-alive/amp/
Granted, no “virus” exists and they are just looking at “viral” RNA which is just as fictitious.
“states that defective viruses may appear similar to exosomes because viruses in the defective state are incomplete viruses with vesicle-like structures.”
This is called a rescue device. They are the exact same particles and can not be distinguished due to the same size, shape, and density. You are using fictional ideas such as “defective viruses” to try and distinguish between the same particles.
“As well, it pertains to exosomes containing and transporting viral parts inside them.”
Again, this is a rescue device. They can not separate these particles in order to be biochemically characterized. This is why they find markers such as “viral” RNA and “viral” proteins in their exosome samples.
“This is because EVs will capture material in a cell, including viruses and/or their parts, and transport them out of the cell.”
This is straight up fiction on your part, unless you care to show where this was seen occurring in a living sample.
“As stated prior, noninfectious viruses (defective) and ‘infectious’ viruses (non-defective) appear completely different in form and are more easily recognized from one another.”
You continue to claim EM images are valid. Please share in any original “virus” study where the particles claimed to be the infectious “virus” were purified and isolated in order to be characterized. This can not happen in a mixed state, otherwise they are just picking whatever particles they want as the representative particle from among millions of similar particles. How is this so hard for you to grasp?
LikeLike
Concerning your other statements:
CPE effects do not determine isolation. It determines the effects a viral sample has on cell tissues when added to them. If those cells show viral replication in and around cells, the sample is claimed infectious to cells. If the virus exists in the centrifuged sample, it ideally should be observable before showing so-called CPE. The traditional “gold standard” used in past years, as you are likely referring to, is no longer relevant to this discussion. Newer technology has begun to replace it long ago.
Crystallography is one such technology. This allows researchers to view structures at the atomic level. You first must have a very pure sample of a virus with little to no debris. This is made possible by utilizing multiple filtrations of a sample, as is described in the isolation manual I referenced in my previous comment which describes the processes in intricate detail (and which you haphazardly dismissed without pause). This sample is suspended in a liquid solution that is then evaporated. The sample evaporates and aligns in a uniform manner that forms crystals. X-rays are then used to defract off the sample, and the data gathered is then used to build full images. This allows the rebuilding and visualization of those structures at atomic resolution.
Each protein of the structure, down to the atomic level, can be inferred and determined in this manner.
I myself have been able to utilize this data to build protein-accurate viral reconstructions for 3D medical animation.
With regard to Hillman:
EM may have caused certain alterations to the appearance of cells, at least in early usage. Likewise, in a similar way, heavy-handed staining used in optical microscopy can cause cells to change some of their structure. Proper usage is required in both situations.
Viruses, on the other hand, even if the nature of EM is detrimental to viral structure in some way, still show an apparent and intelligent structure of the virus itself which proves their existence. No one would rightly infer the exact motion of cellular bodies through EM, but to use it to fill in gaps through static snapshots. Researchers know that cellular bodies appear in a different form in the body itself, but cells can still be seen and observed close to reality. By way of viewing a cell or tissue on a flat surface, such as a slide, you must change from the 3D level as cells would appear in the inner body, to the 2D plane of the slide, thereby causing alterations in cellular movements by nature of the environment. This all must be accounted for.
Things have advanced since Hillman’s time some 50+ years ago with regard to EM and the viewing methods used by researchers (as noted above). EM procedures also advanced in the 40s and 50s, and issues were overcome long before Hillman made his claims. It is also a fact that Hillman was himself claiming non-proven theories that did not have logical sense. He assumed and guessed about what he stated things should look like with little regard for whole cellular processes. He claimed the golgi apparatus, ribosomes, receptors, and other vital cell parts, simply did not exist because he could not see them as he thought they were.
Without the golgi, proteins would float aimlessly in the cell without direction, and lysosomes would not exist, among so much more.
More importantly, all the same so-called ‘artifacts’ he attributed to EM appear under all microscopy techniques. This alone disproves much of his claims. His contemporaries at the time pointed out his errors, saying though there were shortcomings involved in EM, the results observed are close to reality. He refused to heed their advice.
In your world, Hillman can infer characteristics of cells without seeing them, but other researchers cannot.
LikeLiked by 2 people
You have no integrity whatsoever or concern for the truth. You are literally changing the goalposts before everyone’s eyes and gaslighting as you go. Do you think everyone that reads this is that stupid? You sat there and told everyone that the studies in question say something that they do not. When corrected, you ignore that, then go off into your own “rescue device”, which is to claim none of it is real.
I disproved the entirety of your “indistinguishable” claims (and many more) and you simply go off into another tangent because you know the implications if you admit to such grievous error. You most likely do not even realize you are wrong because you are so die-hard.
If you had any honesty or an ounce of self-respect, you would admit you were wrong. But you don’t, and this conversation is over. Continue to spin your wheels, deflect, and deny, as you wish.
LikeLiked by 1 person
“I disproved the entirety of your “indistinguishable” claims”
You did no such thing. You are the only one being disingenuous. Exosomes and “viruses” are only said to be different due to a lack of replication, an unproven function ascribed to “viruses.” Exosomes are said to be INDISTINGUISHABLE from defective “viruses,” which are “VIRUS” particles which can not replicate. In other words, they are the same thing.
“Based on current knowledge, THERE ARE MANY ASPECTS IN WHICH EVs RESEMBLE VIRUSES, IN PARTICULAR RETROVIRUSES. First, although some EVs may be up to a micrometer in size, the majority of EVs are <300 nm, THE SIZE OF A TYPICAL RNA VIRUS. LIKE ENVELOPED VIRUSES, EVs are surrounded by a lipid membrane that also contains cell membrane proteins. LIKE MANY VIRUSES, EVs are formed in the endosomal system or at the plasma membrane via defined biogenesis pathways, for example, involving the endosomal sorting complexes required for transport (ESCRT) machinery (1). LIKE VIRUSES, EVs can bind to the plasma membranes of other cells, enter them either through fusion or endocytosis, and trigger specific reactions from these recipient cells (1). Finally, EVs carry genetic material, and this genetic material can change functions of the recipient cells (2, 3). Especially in the case of retroviruses, EVs generated in infected cells contain selected molecules of viral origin (4) AND CAN BE SO SIMILAR TO NONINFECTIOUS DEFECTIVE VIRUSES THAT HAVE LOST THEIR ABILITY TO REPLICATE THAT THE DIFFERENCE BETWEEN THEM BECOMES BLURRED. In other cases, EVs provide an “envelope” to nonenveloped viruses, e.g., hepatitis A, and these EV-encapsulated viruses can infect cells (5). Similarly, EV released by hepatitis C-infected cells can carry fully infectious viral genomes that in target cells generate new infectious viral particles (6).
In this Perspective, we suggest that in retrovirus infections a variety of diverse vesicles is released, such that on one extreme there are EVs consisting entirely of host cell components and on the other replication-capable viruses. IN BETWEEN THESE EXTREMES ARE NONREPLICATING PARTICLES THAT CAN BE CONSIDERED BOTH AS DEFECTIVE VIRUSES AND AS EVs CONTAINING VARIOUS AMOUNTS OF VIRUS-SPECIFIC MOLECULES (Fig. 1)."
"Moreover, virus infections modify the incorporation of host proteins and RNAs into EVs (light blue). Such infection-induced EVs and the so-called defective viruses and virus-like particles are intermediate entities, AND THE BORDER BETWEEN THEM SEEMS NOT TO EXIST."
"A growing body of evidence indicates that cells infected with enveloped or nonenveloped viruses release EVs that contain viral components. Here, we aimed to create awareness that virus preparations may never be pure but rather are contaminated with diverse subpopulations of EVs, AND SOME OF THESE EVs MAY BE EITHER INDISTINGUISHABLE FROM OR VERY SIMILAR TO SO-CALLED DEFECTIVE VIRUSES. Because of their common biogenesis paths, viruses and EVs may be close relatives, ALTHOUGH ONLY THE FORMER CAN REPLICATE IN CELLS."
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4995926/#!po=0.684932
What is a defective virus?
"A VIRUS PARTICLE that contains insufficient nucleic acid to provide for production of all essential viral components, thus infection is not produced except under certain conditions."
https://www.dictionary.com/browse/defective-virus#:~:text=A%20virus%20particle%20that%20contains,produced%20except%20under%20certain%20conditions.
LikeLike
Welcome, Jeff, though you’re a few weeks or so late if you ask me. That’s when I said that you are mistaking microsomal monooxygenases — the enzymatic nemeses of fat-soluble toxins — for viruses.
What’s your defense? 🙂
LikeLike
Hello reante,
The theory I have proposed is that viruses are cellularly created proteins that are enzymatic in nature, similar to other enzymes but with more specific structures and actions. The class of microsomal monooxygenase enzymes you are referring to are merely fragments of larger bodies and are not whole viral protein structures, nor are they what I claim viruses are.
Viruses and exosomes are larger whole structures and are released from the cytoplasm and the membranes of living or dying cells—whether the cell is healthy, undergoing planned cellular death (apoptosis), or cellular decay. Microsomes are merely parts of a greater whole—they are fragments and cannot be considered whole cellular protein structures. They may be called fragments of larger structures. Therefore, microsomal monooxygenase enzymes are neither exosomes nor viruses.
Viruses are very specific proteins containing mRNA that is tightly woven into their nucleocapsid, which meets with glycoprotein surface receptors on their capsid. This allows for communication between viruses and cells using surface receptor proteins. The glycoproteins (spikes) that cover viruses work by holding a receptive object in place and beginning the breakdown of bonds that hold the structure together through enzymatic hydrolysis, thus causing fractionation of the object. Other enzymes work similarly, but viruses are more unique structures operating on a specific lock-and-key system using mRNA and/or DNA. If this were not true, viruses would begin to dissolve whatever they touch in the body. As with anything in the body, there is intelligence involved.
A virus is a transporter of many enzymes on its capsid. Viruses must be transported safely throughout the body to dissolve specifically, unlike other enzymes, like digestive enzymes that are situated in the intestinal tract.
LikeLiked by 1 person
Jeff, thanks for clearing up my mistake. I apologize. I’ll have to further ruminate on your full comment which you linked to.
What do you mean by the primary exosomal function being as “fluid” carriers? What are these fluids and what are they for? Are you saying they are not nucleic acid- based carriers of information?
LikeLiked by 1 person
You’re welcome… no problem.
With regard to exosomes being fluid carriers, I am referring to them as vesicles that contain components for cellular life; proteins, lipids, nucleic acids, and so on.
LikeLiked by 1 person
Ok right on.
LikeLiked by 1 person
It is true that many toxins are fat-soluble and flow into fat. That is why putting on the proper body fat is essential in our modern-day. The problem arises in people who have very little body fat and low microbial levels due to modern diet. Those without the proper body fat will many times allow toxins to remain free-floating in the blood where they can contact cells and damage cellular life. This is also an issue when cells dump toxins during their cyclical detoxification/cleansing, which occurs throughout the weeks, months, and years of a person’s life as cells renew themselves and their tissues. A skinny body is easily agitated by toxins, and a body that has the proper fat is far less influenced.
Secondly, if a person has very low bacterial levels due to not consuming a diet rich in raw bacteria, their body will not contain the necessary microbes to help consume toxic material and their detoxification symptoms will be far more severe in order to help rid the body of the level of toxins it is having to break down. Regardless, toxic material, whether it is stored in fat or not, must be expelled at some point. To do so, cells make contact with the toxin, such as white blood cells, in order to bind with, escort, and push it out of the body. If the toxin is toxic enough, no living organism, no matter how evolved, will be able to deal rationally with that substance, and it must be taken care of by using a last resort (virus) to diminish the toxin. This is why viral variants exist. Cells alter their proteins to be able to deal with new unrecognizable toxins that are not natural to our bodies.
Lastly, there are many fat-soluble toxins like plastic and heavy metals that are very difficult, if not impossible, for the body to make water-soluble. They embed in fat tissue, cells and their tissue, wherein they settle. To dissolve those, solvents are needed. Sometimes, cells must be fully dissolved to remove the toxin (I.e. polio, heavy metals in the spinal column). This is partly why the ‘infectiousness’ of viruses is an illusion.
LikeLiked by 1 person
>For example, you can easily determine an adenovirus particle from influenza virus particles.
If you’re not careful, this ends up as circular reasoning. “What’s an influenza virus particle? It’s a particle of this shape and size. How do I know its function? It was found in a cell culture containing tissue or fluid from someone who has the flu. How do you know this particle is what caused the flu? It’s an influenza virus. Look at its size and shape. How do you know it’s not an exosome or apoptotic body? Look at its size and shape.”
If you merely label every particle shape X as “influenza virus” and shape Y as “adenovirus” and shape Z as “an exosome,” you haven’t found or characterized anything. You’ve merely given names to recurring patterns of shapes and made stories about their function. Those shapes could turn out to be something entirely different and this method of “characterizing” would never tell you.
>It is mainstream science itself that admits that viruses are able to dissolve the walls of cells.
It sounds like you or whoever started this “viruses=soap” or “viruses=solvent” idea took a mainstream conjecture and ran with it uncritically, and are now wedded to it. TC Fry and Aajonus Vonderplanitz subscribed to a similar notion, so I know it is not new. It’s from back when there were few questioning science as deeply as now, so there was always a tendency to fail to jettison enough of the mainstream paradigm and FUBAR terminology even in the process of radical questioning, apparently on the sentiment that “surely they must be right about *something*” (which is a form of Golden Mean Fallacy, the idea that the truth must be somewhere between two “extremes,” often a sign of seeking social approval or credibility by not coming off as too outlandish).
Since we have seen in every case that their method of gathering information, making inferences, and conducting science in general is flawed at the deepest level, and almost always politically influenced even if indirectly, there is no lower bound that can be placed on the quality of their work.
>The fact remains, that there must be a mediator between cells to deliver cellular fluids and facilitate communication between cells.
Merely because the mainstream makes a groundless conjecture about some intercellular communication object doesn’t mean you jump at it as the means by which cells communicate. That would be just naïve dot connecting as a flailing attempt to make sense of things based on very sparse info and meager debate. It was understandable back in the days before the Internet when debate on such controversial topics was rare to find and made some sense to weave a just-so story as long as it had the right effect (of stopping “virus” fear and vaccination) but the bar has been raised – especially in the last two years.
It’s no longer the Age of Propaganda where the biggest concern is getting the main thrust of the message out there, where being able to say your theory has a mainstream basis helps its long-term viability even when that basis is shaky. With the advent of the Internet, it’s become increasingly important to make one’s arguments ever more watertight. One cannot afford to be right for the wrong reason or within hours they can expect to get skewered on a technicality and the momentum and possibly credibility of the idea lost. The ideas that win are those that are most cleanly correct, thus we have to be rigorous in our approach and not make up stories based on the most-unreliable breadcrumbs dropped by the mainstream.
Take a step back and reconsider whether you really need anything from microbiology to make your case. Fry and Vonderplanitz didn’t have the means or mindset to try to debunk virology at the ground level, so they ran with part of it as that sounds more credible than trying to make the case that “viruses” don’t exist. Now social considerations are less important because information is spread through many anonymous agents. Whatever Cowan, Kaufman, or even Stone might do to harm their own credibility doesn’t appreciably affect the process of refinement of knowledge anymore. The game is now ruthless pursuit of truth, not making compromises for credibility or social standing – or in summary, not making compromises for mainstream palatability, however small.
This doesn’t mean we irrationally throw away everything merely because the mainstream came up with it, but it does mean we have no social hesitation about doing so when it turns out to be based on nothing but seriously faulty processes of discovery. And in general it’s been a very reliable heuristic that the fastest way to make progress is often to deep-six another piece of the mainstream’s paradigm and nomenclature, because they’ve sliced up thingspace to suit their view of how the body works and this slicing up is liable to interfere with our reasoning process in every single sentence we try to utter or reason by until we stop using their terms or at least appropriately bracket them.
LikeLiked by 2 people
>If you merely label every particle shape X as “influenza virus” and shape Y as “adenovirus” and shape Z as “an exosome,” you haven’t found or characterized anything. You’ve merely given names to recurring patterns of shapes and made stories about their function. Those shapes could turn out to be something entirely different and this method of “characterizing” would never tell you.
Nails it!
>Merely because the mainstream makes a groundless conjecture about some intercellular communication object doesn’t mean you jump at it as the means by which cells communicate. That would be just naïve dot connecting as a flailing attempt to make sense of things based on very sparse info and meager debate. It was understandable back in the days before the Internet when debate on such controversial topics was rare to find and made some sense to weave a just-so story as long as it had the right effect (of stopping “virus” fear and vaccination) but the bar has been raised – especially in the last two years
Likewise! Thanks for bringing some … clarity to the discussion. So much of what has been posted in this discussion smacks of “We’re here because we’re here because….” Nothing more than a demand for us to accept the basics of virology because it’s supposedly the accepted “science,” just like the basics of physics, astronomy, never mind that these other sciences, including cellular biology, have the precedence of hundreds if not thousands of years of observations and repeatable experiments serving as a basis, whereas virology (and its outgrowth, allopathic medicime) got its very start as a corporate-sponsored endeavor and engaged in a scorched-earth campaign against all other explanations of related observations, e.g. terrain theory, homeopathy.
LikeLiked by 1 person
Magnificent summary of the state of play.
You da man Big C!
LikeLike
In reply to Clarifire.
Conclusion section of study:
The Role of Extracellular Vesicles as Allies of HIV, HCV and SARS Viruses
The study literally concludes these are different structures from each other, but with similarities. You and Stone are claiming the studies state they are the same when they do not.
The other study being referenced states this: Mission (Almost) Impossible: Separation of Virions from EVs. p.9158 – Extracellular vesicles and viruses: Are they close relatives? Proc Natl Acad Sci U S A. 2016 Aug 16;113(33):9155-61. doi: 10.1073/pnas.1605146113. Epub 2016 Jul 18.
Can you tell me why it matters if you can’t isolate viruses from exosomes if exosomes don’t exist? If viruses don’t exist? Why does it matter? Even if they could be separated with absolute certainty, with absolute guarantee, you would not believe it.
Stone is quoting from two different studies, both of which contain different scenarios.
Either way, it wouldn’t matter if I was cherry-picking the studies or not (and I’m not), neither one of the studies claims that viruses and exosomes cannot be separated, and never uses that terminology, not once. The studies in question, and other studies like them, use phraseology like “almost impossible”, “difficult”, and “challenging”, but not totally impossible. That is Stone attributing things to the studies that do not exist.
Furthermore, there is very likely confusion about what type of viruses they are using in those studies, such as the retrovirus, since the retrovirus may appear more like the exosome, unlike other viruses, which appear less like exosomes. So, this must be understood.
This is being left out by Stone and yourself. If the studies truly meant that viruses and exosomes could not be differentiated, they would not conclude in their conclusion sections that viruses and exosomes are different. They would say that they are exosomes, or that they are all viruses, or that they are something else, but they do not do this. So, that is you and Stone cherry-picking, much more than I ever could.
Everyone can read, once again, it doesn’t matter what I say, what study I provide, you are going to disregard it because it doesn’t fit what you’ve already concluded. So I ask, who is cherry-picking what here?
LikeLiked by 1 person
@Jeff Green
The theme of this blog is to debunk the mainstream using their own sources. It should be clear that a mainstream researcher is never going to say anything greatly unorthodox, so the game is to see if when you take all their claims together it creates problems for the field. Obviously it will be in a way the authors don’t realize because they don’t approach their own field with any criticism of the fundamentals and most scientists also aren’t in the habit of thinking through an entire paradigm as they’re too specialized and compartmentalized; if contradictions and amazing ad hoc coincidences pile up in their storytelling they just assume that’s how things must be down there in the microcosm or assume that’s someone else’s job to verify.
It eventually reaches a level where they would happily contradict themselves or the scientific method within a single paper if it would meet the approval of their peers, just as Enders did when he carried out a control experiment that failed yet nonetheless concluded he had identified the cause of measles, got a Nobel Prize for it, and the rest of the ailing field followed his unscientific methods because “hey, he got a Nobel.”
To a reader willing to question the fundamentals, the problems with the whole theory holding together become starkly obvious *precisely because of and in proportion to* the stark unwillingness of the mainstream to question fundamental assumptions.
When one takes in new information and tries to theorize from it *without* allowing the fundamentals to be called into question at the same time, one is liable to unwittingly resort to ad hocery like “viruses,” “retroviruses,” “exosomes,” and one object containing another as ever more transparent rescue devices to fill in holes in the existing theory. It adds up to a planet-sized violation of Occam’s razor, which is the beating heart of a scientific theory. This is the pattern with all institutional science, though life and social sciences most of all. Physics has its “dark matter” and its ever-expanding zoo of particles and forces for the same reason. It looks really embarrassing and cringey from the perspective of those who understand science and logic, aren’t invested in the dogma of a given field, and aren’t stricken with hyperspecialized “stay in your lane” syndrome.
As to why it matters if “viruses” haven’t been distinguished from “exosomes” if they both haven’t been shown to be any special particles performing their imputed functions, again think of the point of this blog and look at the history of the posts. Virology was resoundingly destroyed on these pages a long time ago, but Mike has continued systematically dissecting and pulverizing each part of its corpse for good measure. This is a great service to the rest of microbiology and serves as a fascinating study of the sociology of scientism.
LikeLiked by 1 person
Clarifire
It was helpful and important that you reviewed the prevailing subculture of this blog as you did, in order to signal to Jeff that he might consider recalibrating his tactics in service of the case he is capably making.
I will add more context. The subculture of this blog is solidly — as in, extremely — in the Deconstructionist movement. Postmodernism was founded on deconstructionism. My senior thesis was on postmodern literature. (In my paper I took a canonical Pomo short story and argued how it was just a phony pomo story dressed up in Pomo style but thematically it was fundamentally modernist lol. I wrote a really good sentence with the word “transmogrification” in it and between that and the accusation of modernism, and probably the fact that it was also in the feminist canon and I am male and opposed to all political co-optation, they hauled me in and accused me of plagiarism.)
Deconstructionism IS the quintessential postmodern act of unlearning the fake Progress narrative of fairytale industrialism, by systematically picking its falsehoods apart but without any care for what’s actually true whether they be any nuggets of truth contained within the falsehoods (remember the definition of evil is truths intentionally placed in service of a falsehood) or, most importantly, holistic truth itself. Deconstructionism and its larger context of postmodernism, are shattered ontologies of a nihilism born of the first-world “spectacular society” of a consumer consciousness in a permanent Disneyland environment. And this is not my opinion. This is postmodernism’s self-acknowledged reality because the point of it for them is to ‘honestly’ acknowledge and explore the nature of contemporary cultural reality as a new, felt ontology, as if that culture is all that exists in the universe. That’s what happens when you live and die in the citadel.
And obviously, Clarifire, you are not a postmodern person, which is why I’m still here. But Mike is Deconstructionist through and through, and he needs to understand that his deconstructionism is as mainstream as it gets because civilization will welcome with open arms anyone who does not care for the holistic truth but merely amplifies the dissonant babble in the great tower, for that is not even a minor threat – it’s precisely the effect that they are looking for.
The idea is to build again in truth and beyond postmodernism. Obviously that’s what Jeff’s about, too, and I’m curious to hear you, Jeff, respond to Clarifire’s possible part-identification of the origins of your paradigm (Frey, Vonderplanitz) as well as continue to elaborate on your own ‘divinations’ about how the intelligent body works and works best, and under what conditions it works best, because that’s ultimately what we’re all here for. Right, Mike?
LikeLiked by 1 person
“The idea is to build again in truth and beyond postmodernism. Obviously that’s what Jeff’s about, too, and I’m curious to hear you, Jeff, respond to Clarifire’s possible part-identification of the origins of your paradigm (Frey, Vonderplanitz) as well as continue to elaborate on your own ‘divinations’ about how the intelligent body works and works best, and under what conditions it works best, because that’s ultimately what we’re all here for. Right, Mike?”
I’m not here to replace one fraudulent theory with another. I am looking for evidence that adheres to logic and the scientific method which does not exist with “viruses” nor exosomes. I do not speculate about the possibilities of particles generated by pseudoscientific means. If you want to engage in fictional discussions, more power to you. It’s a waste of time IMO.
LikeLike
Interesting point about Deconstructionism, reante. I would say Deconstructionism is something applied to culture and tradition, where it throws out the baby with the bathwater. There’s always a “baby” (i.e., something of great value) in culture and [long] tradition, which we discard at our peril.
So one may say the same about the scientific tradition, right? Well yes, we would discard something of great value for those people who work in the field and take money from the public trough. We would throw away their careers and reputations. The mafia has a tradition as well, and I’m sure they discard it at their (their organization’s) peril. But we don’t have to accept it.
Still this is unfair of me; you will say why throw away everything? Surely they did something right. Yes, they surely did something right, but as I remarked before there is no lower bound we can place on how small that something is. Merely because one jettisons almost all of a field doesn’t mean they jettisoned too much. It depends on the field. In particular in medicine frequently the things they do are 180 degrees wrong, seeing things that are wholely beneficial as wholely detrimental. When an edifice is built on going the exactly wrong direction from the start it is hard to salvage much from it, if anything.
Yet I think all this may be a category error: the real tradition that one could apply Deconstructionism to would be the tradition of scientific investigation, and Mike isn’t really doing that. He isn’t saying we shouldn’t have researchers fastidiously documenting observations and performing experiments, and isn’t saying their culture of doing that is lazy. If we look culturally he is actually making a carefully selective critique of the culture itself, though only an implied one: that their culture/tradition has great potential except they need to proceed asocially, ban equivocation, remember to adhere to the scientific method, be more generalist, and have it be perfectly acceptable to question long-held assumptions.
In other words, Mike isn’t deconstructing a culture or tradition; at most he is deconstructing the output of one. Though really he is just applying the most important test that that tradition claims to be built around, and in so doing he seems to be upholding the tradition from back before it went off the rails.
What scientism is really good at is motivating millions of people to get into science who otherwise wouldn’t. Real physics is exciting to me, but it’s not as exciting as the fantasy novel the modern physicists have created and with it inspired a generation. It’s great for setting up the machinery to gather and summarize copious observational data and assemble many potentially useful tools. It’s great for employing many people albeit with net negative effect for the economy. It’s just horrible at theory and therefore horrible at completing the whole process of the scientific method. If all these people’s energy and mostly good data-gathering habits could be directed into something guided by proper theorizing maybe they could do some good.
Now of course if “exosomes” are real and important and the microbiologists are onto something then yes Mike is throwing out the baby with the bathwater, but that is the very question at hand.
LikeLiked by 1 person
Clarifire
Why are dark matter and dark energy so unreasonable? Or are you just saying that the internal logic the establishment uses to justify these concepts is embarrassing?
According to the animist Home Cosmology, source Consciousness and source Energy must exist separately outside of holographic reality/Life. I’ve previously mentioned that gravity is based on Consciousness rather than material/holographic mass; therefore, it stands to Reasin to animism that dark matter exists and they eventually ‘found’ it with their machine eyes because the mysterious gravitational pullings of dark matter pattern to source Consciousness.
And so it is with dark energy outside of the holography; inside, in our reality, Sky the dynamic, pushing, ‘repulsive’ force of chemical bonds in instability that enables the pulling force of Consciousness to animate in creative tension.
LikeLike
On dark matter, reante, the first problem is its clinging to a theory way too hard. As I mentioned, one can always propose an ad hoc fix to explain away a hole in one’s theory, but every such move weakens it (violation of Occam’s razor). It’s much worse when an entity is added to explain just one aspect of a theory and messes up or at least complicates a whole bunch of other aspects.
I think this is another case of grasping at mainstream crumbs just because the mainstream is dominant and the crumbs seem like they might fit with your theory. I think you vastly overestimate the probability that the mainstream has anything much of value to offer.
To be fair, mainstream cosmology, quantum mechanics, and GR are even worse than virology. At least a “virus” is something that could conceivably exist; it’s not a complete nonsensical abuse of language. But an “expanding universe,” “warped space,” etc. are not even meaningful as terms. Modern physicists aren’t even dealing with coherent concepts at the base of the theory.
Dark matter is at least a coherent concept, but it’s invoked to patch up a hole in a belief system built of complete nonsense notions so I wouldn’t pin any hope on it. If your theory independently predicts something like dark matter, I’d just let that stand on its own. At this point the mainstream of modern physics is much more likely to be right about the existence of a theorized entity by sheer coincidence than for any other reason (and of course that too is extremely unlikely).
LikeLiked by 1 person
Control experiments are what these people forgot about science.
The irony is that this happens among those who question viruses too!
Same with Dr Tom Cowan who in his latest video mentions that he is testing orgone devices… He mentioned how he is not scientifically testing devices but avoids an easy test, which I used to test a friend who claimed to have WiFi sensitivity..,
He needs to have someone else use the device or not, and then let him feel if there is any difference from the “action”.
In my case with my friend, I asked her if she felt the WiFi or not, with the router hidden only me knowing if it was on or off. She was as wrong as guessing…. I showed her the data and she refused to believe that it was her thought of WiFi being on, not the actual WiFi that caused the effect.
So if he could do this with orgone, water wand etc, that would be a very good way to filter out the psycho somatic variable…..
And of course we know big pharma doesn’t do that properly as they unblinded the participants 🤑
LikeLike
I don’t think Cowan has forgotten about the importance of controls, even when he talks about or supports “outside the box” topics, like an orgone generator. In that video you mentioned (from July 1st I believe) as well as in many other videos, Cowan is clearly advocating that the scientific method be applied (with controls of course) to all of the seemingly “woo woo” (my term) topics and questions and proposals, such as orgone devices. He says it isn’t an easy task, but possible. He is saying it isn’t happening enough, not that it isn’t needed or should be ignored or applied discriminately.
Indeed, as I interpret Cowan, I see this as one of his central points….that is, that funding and focus etc isn’t applied to studying these matters (no pun intended) because the focus on matter (materialism) is so myopic and thus important “alternative” questions aren’t studied scientifically, they are just ignored out of hand (generally speaking) and so we don’t have the quality studies being done. It’s a systemic issue, I believe he is saying. Cowan is advocating for more, not less, scientific focus on topics that are usually marginalized. He is also saying it won’t be easy, but I would say it’s never easy, which is why methodology in the sciences is such a mess! So, in my opinion, I don’t think it’s fair to imply that Cowan is ignoring or forgetting about controls when in fact in the very video you mentioned he brought up the issue of how to apply the scientific method to these questions. He said he likes being challenged on that point, the very point you are making. He didn’t go into detail, as that wasn’t the focus of the video, but he certainly isn’t saying that controls should be ignored Of course I do agree with your main point about controls being what is forgotten about, amazingly….since as far as I can tell the use of controls are a defining characteristic of what we have been referring to as “science” as all these many decades. I think Cowan applies his own very informal methods (like he said he uses the structured water on different strawberry plants, using controls) while advocating for “real science” to be done in these areas. Can the methods be applied in all areas? I would say obviously not, and that’s where the scientific method is limited, of course. But that’s another topic.
LikeLiked by 1 person
Without adequate controls any result is questionable.
The objective is to prove that the hypothesis can be tested by reproduction of the method stated by the claimant.
If the experiment fails the hypothesis fails. It is why there are postulates like Kochs. Nowhere to date has the existence of a virus been shown. Nowhere has it been proven that a virus causes a disease. They are lying and were getting away with it until people asked questions. It has become very inconvenient for those making the claims.
Good on you Mike.
LikeLiked by 1 person
Well said 👏
It is nice to see people are questioning the evidence presented more and more! 🙂
LikeLike
I am a busy person as I am translating some English language cds into japanese and doing some extra study to prepare for my own version of Spacebusters’ The End of Germ Theory (in japanese of course), but I needed to respond to this:
“Their entire modus operandi is to heavily cherry-pick studies to try to prove their conclusions,”
When the research ALL does the exact same thing, you arent cherry picking. You are using a small sample size to provide examples that people can look into themselves.
And on a side note, I went a step further as a viewer on my bitchute channel asked me to look into veterinary medicine and jabs, I did digging and they do the EXACT SAME THINGS in veterinary medicine as they do for humans: i.e. Bovine Foamy Virus, “isolated” using the EXACT SAME Enders tissue culture method. The same thing for FIV (HIV for cats), which they then use those exact same tissue cultures to make the jabs.
So Jeff, you are EXTREMELY limited in your horizons if you think that ANY of these “isolation” experiments actually isolate anything. I even have a research paper out of Waseda university that connects ALL the dots in a nice little line showing that the whole reason they even use tissue cultures at this point is to make “vaccines,” funded by Takeda Pharmaceuticals (largest drug company in Asia) and the top Science guy at Takeda at the time was yamada tadataka, a former employee of Frazier Pharma and Glaxo-Smith Klein, and the HEAD OF GLOBAL HEALTH at the Bill and Melinda Gates Foundation. The paper OPENLY ADMITS they are testing Vero E6 cells for their use in “vaccines.”
Here is my video on Yamada Tadataka:https://odysee.com/@MaxJenius:7/takeda-yamada:8
And here’s my video on the Waseda paper showing that they admit using these tissue cultures as the basis for jabs:https://odysee.com/@MaxJenius:7/virusesdontexist11:6
LikeLiked by 2 people
Dude,
“When the research ALL does the exact same thing, you arent cherry picking. You are using a small sample size to provide examples that people can look into themselves.”
Then someone else is cherry picking what gets researched. Same difference.
LikeLike
Its not the same difference, and if you cant tell the difference in the nuance between what the quote I responded to was and what you said, you REALLY need to go back and study the English language more.
LikeLiked by 1 person
Dude, I apologize for my careless and incorrect comment.
I will say though that all the research doesn’t do exactly the same thing. That’s bias.nDespite the deep cultural flaws and industrial corruption, they are always trying to refine their processes. For example, Jeff and I are the only people so far who will acknowledge the lengths that the field of electron microscopy will go to improve their freezing methods so that what they look at is the same as what was in the body. (We also recognize the economic boondoggles that these lengths they go to do represent.) Why is it that so few here will acknowledge the degree of vitrification they are now achieving with these samples?
Same goes for ‘clean’ fluid isolation of ‘exosomes.’ Mike’s too proud to let go of the isolation argument in this specific context. He doesn’t need to let go of the functionality (of exosomes) argument because whether to Pattern or not to Pattern: that is the personal question. Nonetheless, Mike makes a religion of the materialistic ‘scientific method’ and so refuses to pattern, and prefers to stonewall those that do reach for patterns because no one can hurt him (his intellect) there.
LikeLiked by 1 person
I’m all for applying logic. If you can logically explain how they can identify the particles claimed to be exosomes in a mixed population of millions of similar/identical particles based on appearance, I’m all ears. Without complete isolation, biochemical characterization is out of the question as the readings could be from any of the substances contained within the sample.
“In these terms, purity means the DEGREE OF FREEDOM FROM NON-VIRAL COMPONENTS, or, conversely, the extent to which viral particles show gross physicochemical homogeneity.”
https://www.google.com/url?sa=t&source=web&rct=j&url=https://link.springer.com/content/pdf/10.1007/978-3-642-85899-4_2.pdf&ved=2ahUKEwi04tPs_7DvAhWOZs0KHZp1BC0QFjANegQIFxAC&usg=AOvVaw0Fx51ebi9lAsov0RyK1wyl
“Because EVs are produced by virtually all cells, probably every viral preparation is in fact a mixture of virions and EVs. To study their respective functions, IT IS NECESSARY TO SEPARATE EVs AND VIRIONS.”
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4995926/
“Still, better and more carefully validated purification methods are NECESSARY TO PREPARE CLEANER VIRIONS AND EV PREPARATIONS BEFORE MANY OF THE PROPOSED BIOLOGICAL FUNCTIONS THAT HAVE BEEN ASSOCIATED WITH EV CAN BE ACCEPTED.”
https://www.google.com/url?sa=t&source=web&rct=j&url=https://www.mdpi.com/1999-4915/12/9/917/pdf&ved=2ahUKEwi8x6SNvcnxAhWGW80KHfVPB3EQFjAMegQIERAC&usg=AOvVaw3kccThKbbHOPZasZ_5KBWb
LikeLike
Very much looking forward to the Japanese version of The End of Germ Theory. It’s a great and rather comprehensive video hitting almost all the right notes with very few missteps, and as you know there is very little material on this in Japanese. Let me know if you need any help with that (though I’ll be unable to through a lot of the summer).
LikeLiked by 1 person
Its cool, i have all the resources here i just need to read through a little more of Poisoned Needle, Private science of Louis pasteur, and maybe some more of Bechamp vs Pasteur. Other than that, I basically have all the important stuff done (as I’ve already translated the drosten paper, and I can explain the Enders method in Japanese well enough for normies).
The only thing im worried about is after passing the idea by a friend who is awakened, she was complaining that 2 hours would be too long. I told her straight up: This is NOT an easy thing to explain and if a person isnt willing to spend the time to actually learn this stuff, because its not something you can just explain, even in the simplest way possible, in under an hour, then they are just lazy. I made what, 10 videos in japanese, all of them around an hour each, going through individual papers and explaining all of the important points, but hearing that even an awakened person doesnt want to sit through that really bothers the hell out of me.
LikeLike
Could make a condensed version for the lazy. But really it’s probably that they’re not expecting the level of complete obliteration of virology that the video provides, because again the standard of info in Japan on this stuff is almost all weak sauce.
LikeLike
Well, in conclusion, the only purpose of germ theory and the accompying invention of virology is to promote disease and illness. The result of that is the endless marketing of big pharma drugs, vaccines and now mRNA gene therapy injections as being the only things that can save you from an early grave. In truth, those are the substances more likely to kill you off sooner.
LikeLiked by 1 person
Exactly 💯
LikeLike
Mike, I didn’t realize you now believe in genomics and the possibilities of gene therapy.
LikeLiked by 1 person
How does this mean I believe in genomics and gene therapy? Just because they claim something contains mRNA and is a gene therapy does not make it so. The injections are nothing but toxic concoctions.
LikeLike
Because you replied “exactly 100pc.” I’m just noting the intellectual slop is all like George with the grammar.
LikeLike
Yes, it is claimed that the injections contain mRNA and that they are gene therapy. That is the claim. I agree that is the claim. That does not mean that I agree that this is what the injections are composed of nor how they act within the body.
LikeLike
nestorseven called them gene therapy injections without qualification. You agreed 100pc. Case closed mike
LikeLike
“The result of that is the endless marketing of big pharma drugs, vaccines and now mRNA gene therapy injections as being the only things that can save you from an early grave.”
My interpretation is that he was talking about how things are marketed to save us. If you think this is a “win” for you based on your interpretation, so be it. Obviously, I do not agree with mRNA and gene therapy. I agree it is being marketed that way as a solution.
LikeLike
Marketing, unqualified and in and of itself, doesn’t mean lying/bullshitting. Really we would need to say, “shit marketed as shinola” in order to grammatically justify your interpretation, which would no longer be an interpretation but instead a correct reading.
But I see you’re interpretation as stated and that that may in fact be what nestorseven meant I have no idea.
LikeLike
In reality, regarding the microscopic level of existence, no human knows what happens in a living organism. To believe that optical or electron microscopy reveals what exists and what happens in the living organism is madness in its purest form. Molecular biology, genetics, virology, immunology, etc … all these are just pseudo-scientific tales about things never proven to exist.
LikeLiked by 1 person
What so-called microscopic scientists do at the microscopic level is insanely absurd, ridiculous, and manipulative. There is no trace of science in what is done in so-called molecular biology laboratories. Everything is a colossal scam generated by the madness of vanity in the pursuit of money and glory.
LikeLiked by 1 person
Earth to Nike, light microscopes are magnifying glasses.
LikeLiked by 1 person
If you want to understand what human science is, especially in areas that go beyond visible evidence … read the fairy tale about the emperor’s invisible clothes, written by Hans Christian Andersen.
LikeLiked by 2 people
By that measure Nike you must just be talking to yourself and listening to yourself everytime you visit the Viroliegy comment section.
LikeLiked by 1 person
Thanks! I will check it out. 🙂
LikeLike
That tale is indeed very telling! I want to someday read all the versions and maybe write about the insights, it’s a very deep allegory.
LikeLiked by 1 person
Now this is where the rubber meets the road. Again there is no proof of transmission or replication of any so-called virus particle. All we have is a bunch of debris from dead cells and unproven theories. This is great work.
Mike, you should back it up and keep copies just in case.
Not to be picky, but there might be a few grammatical problems – maybe. See what you think.
other’s noticed = others noticed
The cellular debris, as exosomes were originally claimed to be, has (= have) been picked as representatives for many fictional entities including “viruses” and exosomes.
LikeLiked by 1 person
Thanks again George! I always appreciate grammar corrections. I try to catch them all but I am not always successful. Not making excuses, but I have a demon phone that constantly changes things I spell correctly into non-existent words. For example, it will change “that” into “thar” and “but” into “bur.” It constantly changes “particles” into “partickes.” I thought these were my mistakes until I saw my backstabbing phone switch them before my very eyes. Technology…can’t live with it, can’t live without it. 😉
LikeLike
Lol! I still cannot believe you write this entire blog on your phone, and a demon phone at that!!!
LikeLiked by 1 person
Lol, yeah. I’m a glutton for punishment 😉
LikeLike
George I notice that your use of exosomes is free-standing without quotes around it yet they accompanied viruses. I must be making progress against all odds. 😉
LikeLike
Reante,
“Matter can change form through physical and chemical changes, but through any of these changes matter is conserved. The same amount of matter exists before and after the change—none is created or destroyed. This concept is called the Law of Conservation of Mass.”
If this law is true then what do you want to call the breakdown products that result from cell death?
LikeLiked by 1 person
I’m not sure what you’re getting at George so feel free to let me know if that seems useful, but fundamentally I would call a dead body (“breakdown products”) food. Apoptotic bodies are food for saprophytes. Jeff’s recent comment to me about the body directly recycling this food to any appreciable degree, for it’s own use, does not appear to stand to Reason to me because I don’t see a mechanism for it. If Jeff’s reading this comment I’d love to hear about the insides of that claim. Seems to me there are highly likely to be some metabolites from saprophytes eating our dead tissues that we may be to put to use but calling this any kind of recycling program would be a stretch; the saprophytes recycling our death into their lives. We grow cells anew, by cell division, and we fuel that by working our eating drinking breathing and sunning faculties.
LikeLike
In reality, True Medicine has nothing to do with the false “sciences” in the field of microscopy. All the religions of the world gathered together are not able to emit 0.1% of the lies and errors of all branches of Western Medicine. But the most prolific branch of Modern Medicine in terms of lies and errors is the one based on optical microscopy and, especially, on the so-called electron microscopy. The number of lies issued by all branches of so-called molecular biology tends to infinity if we add to it the lies in the field of so-called chemistry.
LikeLiked by 1 person
Right Nike, and there’s a wall of ice surrounding the flat earth and nobody’s. Western Medicine’s lies are fleas on the back of the dog that is religion. The flea cannot be born without the dog, and the flea infestation (western medical totalitarianism) on the diseased dog is a symptom of a supremely rotten Judeo-Christian religious dominance.
Which makes you the germ theorist placing the blame on the fleas that correlate with symptom rather than recognizing the ecological conditions of the terrain as being the causal factors.
LikeLike
How about this, son….with technology, I can prove conclusively that there is no curvature (e.g. ever heard of a Nikon P900), but you can’t prove the same…why is that, son?
LikeLike
Potente, you’re living inside a parlor trick because it helps you feel like the world can’t hurt you as much from in there. You need to man up and go outside and earn yourself a real living.
LikeLike
Reante, please refrain from put-downs. If you can’t debate without engaging in logical fallacies, then don’t comment.
LikeLike
Loud and clear boss. Looks like I might be drawing closer to being your first after all. 🙂
LikeLike
There’s no reason to be. We just need to keep discussion civil and focused on the evidence. There is no need to take cheap shots.
LikeLike
I thought it was an accurate diagnosis and sound advice myself, for a flat-earth adverse reaction.
LikeLike
It was an ad hominem attack. Let’s skip the logical fallacies and not make things personal.
LikeLike
Mike, as a “ghost writer” myself, I’m pretty sure “reante” and “Jeff Green” is the same person. While they attack from different perspectives, they both exhibit the same aloof condescension. I would guess that he probably works for a PR firm contracted by Big Pharma. If your blog gets bigger, he’ll get help from others. I wouldn’t be surprised if he’s also “Potente Angelo.” He creates a superstitious strawman and then slays him with his reason.
LikeLike
While you could be correct about Reante and Jeff Green, I know for a fact Potente is not related to Reante. I’ve known Potente for a while and he is very much against all of this. I won’t be surprised if more people chime in trying to discredit the information here. Won’t be the first nor the last time. 😉
LikeLike
Only a POS ghostwrites. Still waiting on Jeff to address Clarifire’s patterning of his outlier ‘virus’ theory to Vonderplanitz and Fry.
LikeLike
That’s a truly pathetic and paranoid low blow, Mike.
LikeLike
What is a “truly pathetic and paranoid low blow?”
LikeLike
Concurring that Jeff and I could be the same paid plant.
LikeLike
I’m not sure how you think I could rule the possibility out as I do not personally know either you or Jeff. Do I think you are the same person? No. In all honesty, Michael S and Jeff had more similarities in their writing styles but I don’t think they are the same person either. Can I rule out the possibility that all of you or none of you are ghost writers? No. There are only a few handful of people who comment here that I know for sure are not. Potente is one of those.
LikeLike
Where can I redpill myself on chemistry? I’ve been meaning to study dissident chemists, if there are any.
LikeLiked by 1 person
I saw some YT videos by That Chemist on falsified chemistry data… that may point you in the right direction.
LikeLiked by 2 people
My oh my, Mike, you hit some people’s nerves! A sign you are on to something. 🙂 Thanks again.
LikeLiked by 3 people
Lol, I see that. 😂 Who knew so many needed replacing one fictional entity with another? 🤷♂️
LikeLike
So many people need to have a bogeyman to blame stuff on. 🙂
LikeLiked by 1 person
Or a replacement theory. They believe that the questioning the dogma must provide a better alternative which is absolutely ridiculous.
LikeLike
A little information about adeno”viruses,” which have allegedly been used to make viral vector vaccines like the notorious AstraZeneca covid injection. This is how they say it works, but proof is lacking.
“Adenovirus-based SARS-CoV-2 vaccine
A coronavirus vaccine known as ChAdOx1 nCoV-19 or AZD1222 was developed by the University of Oxford and AstraZeneca to treat severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) infection (the cause of COVID-19). In this vaccine, a modified version of a chimpanzee adenovirus (ChAdOx1) is used which can enter human cells but not replicate inside. A gene for the coronavirus vaccine was added into the adenovirus DNA, allowing the vaccine to target the spike proteins that SARS-CoV-2 uses to enter human cells.”
https://www.news-medical.net/health/What-are-Adenovirus-Based-Vaccines.aspx
“Introductory Chapter: Human Adenoviruses”
This covers just about anything you might ask about Adenovirus.
https://www.intechopen.com/chapters/64829
“Isolation of a variant human adenovirus identified based on phylogenetic analysis during an outbreak of acute keratoconjunctivitis in Chennai”
This is a highly detailed paper on how an adenovirus was isolated (as defined by virologists). It defines the process with remarkable accuracy.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3461738/
“Human adenovirus binding to host cell receptors: a structural view”
How they think adenoviruses work.
https://link.springer.com/article/10.1007/s00430-019-00645-2
Virus Structure
“Examples of viruses with an icosahedral structure are the poliovirus, rhinovirus, and adenovirus.”
https://morgridge.org/community/teaching-learning/virology-immunology/factsheets/virus-structure/#:~:text=Viruses%20with%20icosahedral%20structures%20are,poliovirus%2C%20rhinovirus%2C%20and%20adenovirus.
Some icosahedral viruses look a little spikey like the SARS-CoV-2 virus
https://pubmed.ncbi.nlm.nih.gov/10585969/
“Emerging role and therapeutic application of exosome in hepatitis virus infection and associated diseases”
“Exosome mediates HBV replication and transition”
At some point hepatitis B virus core particle is a icosahedral object.
“Subsequently, HBV core particle is completely packaged into an icosahedral capsid to form a replication intermediate sphere with a diameter of 42 nm, then being released out from the original infected cells [32].
https://link.springer.com/article/10.1007/s00535-021-01765-4
So why aren’t these icosahedral objects simply a certain kind of exosome?
“Exosomes cloak the virion to transmit Enterovirus 71 non-lytically”
So the exosome is a cloaking device now.
“Enterovirus 71 (EV71) is a non-enveloped virus and it can be released from host cells through a traditional cytolytic manner. Now, we showed EV71 could be spread non-lytically between cells during early viral infection. In order to explain this phenomenon, we separated supernatant fluids of rhabdomyosarcoma (RD) cells cultures infected with EV71 by isopycnic gradient centrifugation. Two populations of virus particles were morphology indistinguishable by transmission electron microscope (TEM). It showed that some EV71 particles were wrapped inside extracellular vesicles which were verified to be exosomes by immunoassay and morphologic analysis. In addition, exosomes containing viral RNA were shed in plasma of EV71-infected encephalitis in children. Our findings indicate that the “non-enveloped” EV71 virions could be wrapped within exosomes which promote their spread in the absence of cell lysis.”
Introduction
‘EV71 is a positive single-stranded RNA virus that belongs to the family Picornaviridae, genus Enterovirus, species Enterovirus A. The EV71 particle is symmetrical with a 20–30 nm icosahedral capsid.’
If the particles were morphology indistinguishable then are some exosomes icosahedral or no?
https://www.tandfonline.com/doi/full/10.1080/21505594.2019.1705022
LikeLike
Mike, please elaborate, is there someone on this comment thread claiming viruses cause illness? If so, can you point these people out to me?
LikeLike
I don’t believe Jeff says “viruses” cause illness. However, he believes “viruses” exist and that they act as soaps/solvents in the body. In other words, he rejected the mainstream unproven pathological functions assigned to these fictional entities and assigned other unproven functions on the same fictional entities. 🤷♂️
LikeLike
OK, you just let me know when someone comes to these threads and claims viruses are pathogens….my financial backers have a proposition for ANYONE who makes these claims…
LikeLiked by 1 person
I will definitely send them your way. 😉
LikeLike
This is probably what he is talking about. The question is can this process be seen with optical microscopy at least in part?
“In human adults, billions of cells die every day as part of the body’s natural processes. Cells that become damaged by microbial infection or mechanical stress also die. The cell death that occurs in the physiological setting is programmed, and is therefore called programmed cell death (Lockshin and Zakeri, 2001). Apoptosis is the major death process, but necrosis and autophagic cell death have also been proposed to play roles in programmed cell death (Kroemer et al., 2009). Dying cells secrete a “find me” signal, and they expose an “eat me” signal on their surface. In response to the “find me” signal, macrophages approach the dead cells; they then recognize the “eat me” signal (Ravichandran and Lorenz, 2007). Using sophisticated cell machinery, the phagocytes ingest the dead cells, direct them to lysosomes, and degrade their cellular components into basic biochemical building blocks: amino acids, nucleotides, fatty acids, and monosaccharides. These molecules will be released from the lysosomes and reused to make new macromolecules. In definitive erythropoiesis, the process by which red blood cells are generated, the nuclei are extruded from erythroid precursor cells at the final differentiation stage and are engulfed by macrophages (Chasis and Mohandas, 2008). The machinery used for the engulfment and degradation of the extruded nuclei appears similar to that used for the removal of apoptotic cells.”
https://www.sciencedirect.com/science/article/pii/S0092867410001297
LikeLiked by 2 people
From my understanding, this process can not be observed unless through EM, i.e. static images of dead samples.
LikeLike
There are videos of macrophages eating things.
LikeLike
Nice find George. Like you say this is presumably the mechanism Jeff is talking about. It does appear to be a recycling program I wasn’t aware of but as I recall Jeff was talking about recycling in the context of some level of inflammatory disease which would require saprophytic activity due to the sheer numbers of dead cells requiring decomposition. Here’s an article talking about the interface between the body’s recyclers and saprophytes, with the saprophytes of course being misrepresented/mischaracterized:
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4987732/
I like the phrase they use – “apoptotic corpse.”
LikeLike
Way too many variables to consider in that article. I am more interested in how they came to their conclusions rather than in the conclusions themselves. In other words, show me the evidence and explain how it was obtained. Nevertheless, I don’t think they proved the existence of transmissible disease causing particles that enter cells, highjack cell machinery, replicate and kill the cells in the process.
Additionally, I always like to know if the authors are evolutionists because they look at things as evolving instead of created. It is easy for them to believe that numerous types of viruses are lurking everywhere just waiting for an opportunity to enter human cells by means of receptors that match them perfectly. And then afterwards an immune system sends an army to counter attack in a specific way because it has been trained to do so by previous engagements. Why not avoid the whole mess and just not create viruses in the first place?
LikeLiked by 1 person
Some say that the germs (if that’s what you want to call them) or the cleanup crew can become toxic themselves if they are deprived of oxygen and die. Just something else to consider. If this scenario or something like it is true then what could possibly happen if non-biodegradable elements are encountered? Take for example heavy metals that make up the adjuvants in vaccines and graphene oxide nanoparticles and other types of nanoparticles.
LikeLiked by 1 person
@reante
Do you really mean that? 😉 I think you’d prefer to gesture at counterarguments rather than spell them out, and that is fine as not everything true can be readily articulated and often the other party will get an idea from a hint, but I don’t think an aggressive style couples well with that approach.
By the way, I’ve been thinking about your idea of animism and it occurs to me that isolated tribes will be in much more intuitive touch with the state of things and probably also have healthier mental hardware than us, so there’s no reason to discard their “religions” or beliefs as primitive. There technology is primitive but technology is not science nor is it understanding of how the natural world works.
Moreover, these tribes are not blinded by Western reductionism and as such may be more far suited to the study of nature than we are. Indeed our understanding of natural systems (emergent orders / spontaneous orders) can be called primitive. Even F.A. Hayek did only the most broad-strokes analysis of them (and limited his study to emergent orders of human interaction).
The bad translations from tribal language, e.g., into “spirit” instead of “consciousness,” and the philosophical conundrum presented by the mind-body problem (i.e., if consciousness is only a chemical reaction, why do *I* feel so subjectively like an experiencing entity?). And also what happens when we die. It had occurred to me that the one answer that does work is to say all matter has consciousness and there is nothing but matter. Perhaps the consciousness of a plank of wood or a plastic bag is very unidirectional or something. In other words, I hold in a high place in my provisional thinking the hypothesis of animism – the religion of the most (technologically) primitive.
As for connecting this to science, as I said I think there is too much grasping at false leads that are seen in the mainstream. I’m pretty sure that reductionism and the study of natural systems are two different paths and never the twain shall meet. Naturally I can only await a counterexample, and maybe you or Jeff have one you’re willing to spell out here in detail rather than just give hints, but long experience with the (seemingly) inevitable result of such pursuit has reduced my eagerness to spend a lot of my own time on such claims.
LikeLike
Hi Mike, Great work again. Please consider as one of your next topics the cell theory of biology. I’m sure you have heard Dr. Cowan talk about this theory also being misunderstood and maybe even completely false, especially in the light of Harold Hillman’s work. ViroLIEgy relies heavily on cells, receptors etc. and if all that is false then most of medicine and biology comes tumbling down. Keep up the great work, George Hilgendorf
LikeLiked by 1 person
Thanks George! I definitely plan to tackle cell theory. It’s on my list! 😉
LikeLike
Exosomes to the rescue
Exosomes decorated with a recombinant SARS-CoV-2 receptor-binding domain as an inhalable COVID-19 vaccine
“The first two mRNA vaccines against infection by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) that were approved by regulators require a cold chain and were designed to elicit systemic immunity via intramuscular injection. Here we report the design and preclinical testing of an inhalable virus-like-particle as a COVID-19 vaccine that, after lyophilisation, is stable at room temperature for over three months. The vaccine consists of a recombinant SARS-CoV-2 receptor-binding domain (RBD) conjugated to lung-derived exosomes which, with respect to liposomes, enhance the retention of the RBD in both the mucus-lined respiratory airway and in lung parenchyma. . .
. . . Virus-like particles (VLPs) and nanoparticles (NPs) are powerful drug-delivery carriers16. In particular, exosomes are a type of naturally occurring extracellular vesicle found in the body, which makes them a native and ideal delivery vesicle for targeted drug delivery17,18. Because they carry and express their parent cell’s RNAs, proteins and lipids, and because they express surface proteins and receptors from the parent cell, they are superior at targeting same tissue-recipient cells19,20. They contain a cocktail of molecular components composed of proteins, lipids and nucleic acids with therapeutic properties21. Furthermore, exosomes can be engineered by creating surface modifications to express proteins or peptides to enhance targeting19,22.”
So what’s the difference between a viral particle and an exosome?
https://www.nature.com/articles/s41551-022-00902-5
LikeLike
Certainly there is something beyond what we can see through the most powerful optical microscopes. But so-called indirect evidence does not have the intrinsic power to help us get past the stage of assumptions about what exists and what happens in the sub-microscopic realm. The so-called indirect evidence only helps us to understand that the structuring of existence goes beyond the field of visibility but without giving us any concrete evidence to know how to structure energy existence beyond the visible spectrum and what happens in the sub-microscopic field. The truth is that in its attempt to know existential reality, Humanity cannot exceed the limit of visibility, which is why all the so-called structures purported to be sub-microscopic are nothing but assumptions never directly and uninterpretably proven.
As for the technical-scientific ways in which so-called scientists obtain so-called indirect evidence, the vast majority of them are highly questionable. In fact, the so-called human science is made up of close to 100% only of assumptions presented as truths, based on the so-called scientific authority, the so-called consensus of scientists, the so-called reference laboratories and the so-called peer review process.
In reality, there is no science in the so-called science of humans. There is only a constant effort to understand existence, which is embodied in billions upon billions of tests and trials on the basis of which so-called scientists try, without much success, to understand what exists and what is happening. The problem is that the whole system educates the masses to blindly believe in the professional skills and honesty of so-called scientists, which is why almost no one doubts anything about so-called science.
As for the so-called electron microscopy, a well-known so-called electron microscopy specialist said that it does not matter so much what shadows and lights appear in black and white micrographs, but what interpretations are given to the shadows and lights that appear in micrographs. So, although so-called electron microscopy is nothing more than a game of assumptions, people blindly believe that so-called electron microscopy is an exact science, although not even the creators of so-called electron microscopes know anything about how laboratory preparatory techniques and electron microscopy itself modifies the inorganic and organic matter under observation because they do not have any comparative pattern obtained by optical microscopy. Like so-called nucleic acid purification and multiplication procedures, electron microscopy is nothing more than another scam of religion called scientism.
The truth is that the so-called scientific world does not have any capabilities to be able to see in real time and without any negative influence, what exists and what happens in the sub-microscopic field in the living organism. Almost all so-called submicroscopic science consists of assumptions imposed as truths through manipulation, money, and coercion.
LikeLiked by 1 person
About Rife and his microscopes. I’ve been studying this information on and off for a while now and I still don’t know what to make of it. The problem is to understand what he did to overcome the diffraction limitation of light and achieve the magnifications he claimed to have obtained. In any event the man certainly had the background and the training to make an attempt to do it.
“Dr. Royal Raymond Rife, Jr., Ph.D. was born in Elkhorn, Nebraska on May 16, 1888. His mother died when he was just eight months old. Because his father, a mechanical engineer, worked 14-16 hours a day, the young Rife was put in the care of his aunt, Nina Colber Rife Dryden, who raised him as her own.
In 1905, he entered John Hopkins University to pursue medical studies, but his interest in bacteriology took him into the world of microbiology. He then attended Heidelberg University in Germany, where he developed photomicrographs for their Atlas of Parasites. Heidelberg University was so appreciative, they awarded him an honorary Doctor of Parasitology.
From 1904 to 1908, Rife worked with Hans Luckel, Carl Zeiss’ optical scientist and researcher at Zeiss Optical Works. Passionate about the ability of microscopes to make organisms visible to the human eye, Rife set out to improve their resolution and magnification. . .”
https://www.biologicalmedicineinstitute.com/royal-raymond-rife
LikeLike
Why couldn’t one use Microsphere Optical Microscope to image a living virus? It has resolution down to 80nm for the current gen and the next gen will be at 60 nm (https://cde.nus.edu.sg/ece/news/phaos-technology-launches-disruptive-microsphere%E2%80%90assisted-microscopy-technology-in-collaboration-with-sigmakoki/). https://www.labcompare.com/49-Digital-Microscope-Video-Microscope/14241786-NANORO-M-Materials-Microsphere-Nanoscope/
LikeLike
If I’m reading your link correctly, each virus would be represented by two to five pixels. 1080 resolution is 1,920,000 pixels per frame. Imagine trying to recognize a person at 1/1,000,000 of your TV screen.
LikeLiked by 1 person
From the comments:
– Daniel Roytas
Do you know if there is an imaging technique that can see a liver cell, or a brain cell, or a kidney cell in a living human being?
– Robert Griffis
The soft tissues of living organisms, such as human beings, are made up of water crystals and mitochondria, or what Antoine Béchamp called microzyma. When organisms die, or their tissues die, the water crystals form what appear to be cells. Harold Hillman exposed the cell fallacy around 1980 or so, but that hasn’t stopped all of the nonsense about “cells.”
– Robert Griffis
I’ve heard the cell fallacy called a “death cult.” As in, there are cells in dead, dehydrated tissues, but never in tissues of a living organism. Skin cells in the surface of your body are already dead. Blood “cells” I imagine are only cells when separated from a living organism. Blood, milk, and other bodily fluids are very different outside of the body than inside.
– Robert Griffis
One of the most common errors in science is to develop a rigorous method of observation, but in so doing, to completely lose sight of the object of study. The object here being health and disease in living organisms. So much of science based on modern microscopy is total 🐂💩.
Physicists and virologists making up all kinds of stories of what they “see” under a microscope. Not understanding what is done to samples to make them visible. This is what Hillman is referring to as cellular biology mostly being artefacts.
And yet these artefacts become a part of widely accepted “science.” I flinch anytime people mention “cells” or “viruses.” So I think it’s good to get this cell nonsense out of the way. I prefer to refer to parts of living organisms as tissues.
The cell delusion may be even deeper than the viral delusion. We’ve made up entirely false paradigms, that have become universally accepted mythology that passes for science. We need to correct that, or we’ll be hopelessly lost.
LikeLiked by 1 person
I’m curious, if blood cells die when removed from the body, how do blood transfusions work? Possibly the body can somehow recycle the dead material, but still. I thought there were videos of blood cells flowing in the body, but maybe I’m just recalling CG.
LikeLiked by 1 person
I think Nike may be a timely flanking maneuver by Mike lol. Or is it a rearguard action? Just kidding Nike. I mean Mike.
Noticed this statement from you Mike in your clash with Jeff: “They can not separate these particles in order to be biochemically characterized.” Didn’t know you were getting into farming. I really wouldn’t recommend line breeding, though – it might become profitable. Just let the acids do they’re own thing, come what may.
LikeLike
Via Daniel Roytas
Is there a case in the scientific literature, where a cell and all of its organelles, has been observed, whilst connected to a living host?
If we are to be truly scientific, we must eliminate the process of removing tissue from a living host as a potential extraneous variable.
The definition of an extraneous variable is as follows – “In an experiment, an extraneous variable is any variable that you’re not investigating that can potentially affect the outcomes of your research study. If left uncontrolled, extraneous variables can lead to inaccurate conclusions about the relationship between independent and dependent variables”.
In this particular circumstance, removing the tissue is the extraneous variable. Where is the experiment that has proven that removing tissue from a living host does not impact the tissue sample we are seeing under the microscope?
In order to do this, a cell and all of its organelles, must be shown to exist (in the same way when detached from the body) before it has been removed from the living host.
Otherwise we are simply pointing to the effect, without any understanding of the cause.
We need a baseline or a validation experiment in order to be able to draw any conclusion. If we do not have it, we are being anti-scientific.
This is a case of affirming the consequent logical fallacy.
LikeLiked by 1 person
Daniel Roytas is quite right. It really means cellular biology and much of physiology at the small scale is not science and perhaps never will be.
They will say “we’re doing the best we can” with their highly unscientific guesswork, but by presenting that guesswork as established fact they do unimaginable damage to the world (well, it’s imaginable because we’re experiencing it right now).
There are much better ways of approaching health, physiology, and agriculture. These involve natural orders optimizing for survival and harmony so they can be studied by rational agent-based reasoning, just as we study another human being’s motivations and communicative considerations.
This is how we end up with ideas like influenza being a deep cleaning the body performs in coordination with you (whatever timing considerations you have for your life convenience) and other members of your tribe. It also provides a theory of fasting, a crucial piece of the disease puzzle that I hope to elaborate on subsequently.
LikeLiked by 1 person
Institutionalized organizations can’t do true science because of the organizational stupidity that is axiomatic to all institutions, due to vested interests. All organization gets institutionalized beyond Dunbar’s Number.
The word ‘science’ should be dropped altogether. It’s just a relativistic bastardization of the fruits of the cumulative and accurately patterned observations on cause and effect in the local ecology.
LikeLike
“Institutionalized organizations can’t do true science because of the organizational stupidity that is axiomatic to all institutions, due to vested interests. All organization gets institutionalized beyond Dunbar’s Number.
The word ‘science’ should be dropped altogether. It’s just a relativistic bastardization of the fruits of the cumulative and accurately patterned observations on cause and effect in the local ecology.”
Agreed on all of this, reante, though science is possible in non-living systems where variables can be isolated. Institutions are never going to be reliable even on that front, though they can catalog useful observational and experimental data.
The word “science” should be limited to fields where variable isolation is possible and theorizing about unobserved mechanisms is necessary. Basically the “hard” physical sciences.
Mathematics, Austrian economics, our various reason-based arguments about physiology, and other such investigations have been called “science” by many in the past (e.g., Mises called praxeology the “science of human action”), but a word like “logic” or “rigorous reasoning” or “naturalistic reasoning” or “emergent systems analysis” should suffice. There is absolutely no implication that these fields are less important than the sciences. Trying to ape the sciences by “upgrading” these fields to “real sciences” was huge mistake that has made modern humans sick in so many ways. As I say, reductionism and natural orders never mix.
LikeLiked by 2 people
This technique provides some fascinating imaging. Interpreting it is another matter.
Cutting-edge microscope spies on living cells inside the body
Imaging technique captures 3D video of cells at work in unprecedented detail.
“A microscope that combines two imaging techniques — including one used by astronomers — now allows researchers to capture 3D videos of living cells inside organisms. . .”
https://www.nature.com/articles/d41586-018-04760-7
LikeLike
Yeah nice find George. Looks like energy and that which animates it.
As above so below.
LikeLike
Because some have researched more deeply and started exposing the scam of using optical microscopy and, especially, the scam of using electron microscopy in the field of biological research … now, scammers of the so-called science have invented another scam in the field of microscopic imaging which , unfortunately, it is unconditionally believed by 99, (9)% of people, just as the results of optical and electron microscopy techniques were unconditionally believed.
LikeLiked by 1 person
In reality, blood transfusions are not at all beneficial and do not work at all. The scam with blood transfusions is much simpler than it seems (in fact, it is identical to the scam of using synthetic drugs and vaccines). So, if a person has enough vitality to survive and, especially, if he has enough vitality to heal after a transfusion of whole blood or primary blood components, doctors will declare that the transfusion has worked and that survival and healing is due to transfusion. If the patient dies after transfusions of whole blood or primary blood components, doctors will claim that the trauma or illness was too advanced to do anything.
LikeLiked by 1 person
What about dialysis Nike? How does ‘all’ the blood get removed from the body, ‘cleaned,’ and put back in the body twice a week for decades sometimes, if blood immediately changes deleteriously and significantly outside of the body? The answer is it obviously doesn’t.
LikeLike
Hmm, Nike that’s interesting on blood transfusions. One less reason to ever set foot in a hospital if true. It would be great to be able to discard even more of mainstream trauma care, which looks quite solid in many ways.
LikeLike
Dialysis is not an allogeneic blood transfusion. During dialysis the blood is removed from the body gradually and for a short time. This does not mean that the dialyzed blood does not suffer any degenerative changes but that the degradation generated by the dialysis process is small enough so that the dialysis patient’s body can regenerate it. By courageously and objectively analyzing transfusion and dialysis, anyone seeking the truth will understand that the two procedures of modern Western medicine are nothing more than criminal scams.
LikeLike
Western medicine promotes an endless array of criminal medical deceptions using fear manipulation. So instead of identifying and eliminating or reducing the harmful factors that force the body to adapt, altering their tissues and functions in order to survive, so-called modern Western medical scientists are resorting to criminal medical scams such as tissue and organ transplantation, transfusions of whole blood or primary blood components and dialysis. In reality, the so-called Western doctors do not understand how our being works.
LikeLike
Nike, I can understand blood transfusions and dialysis possibly being scams, but how would you explain things like heart or kidney transplants?
LikeLike
Nike the “harmful factors” are structural to civilization. Allopathic medicine is the dominant paradigm exactly because both the inner and outer terrain has become quite fouled for the vast majority of people. Allopathic medicine is basically palliative care. That’s why Tulsi Gabbard called it the sick care system at the Democratic debate.
The medical scam is a function of the civilizational scam. Rats in the capitalist rat race don’t have much say over whether or not their terrain is healthy, therefore palliative care — symptom suppression (and for-profit) — is the only available option.
LikeLike
Yes reante it’s a consequence of wildly exceeding Dunbar’s number. Medicine as a cultural phenomenon is about squeezing the rats to keep them in the race 9-5. Low symptom expressiveness is the name of the game. Same reason the Fed lowers interest rates whenever the market tanks. And overaggressive firefighting. Trying to delay the detox and squeeze more out of the rats so they stay in the race and never duck out to actually allow their body (or the economy, or the forest) to cleanse and heal.
It’s palliative care for everything, with any “science” stories needed to prop up the system, but as individuals we can just opt out.
LikeLiked by 1 person
The more bombastic a medical deception, the stronger it is unconditionally believed. This is also the case with transplantology. Almost no one can accept that there is even a small possibility that transplantology is still a colossal criminal scam in Western Medicine. People simply refuse to believe that they could be so deeply deceived. People can’t believe that transplantology is just a lie, just smoke and mirrors. And precisely because of this attitude of the people, it is meaningless to discuss this subject by asking questions that would revolt them, because they cannot accept that doctors would be willing to commit such criminal scams through so-called tissue transplants and of allogeneic organs.
LikeLike
Well Nike, it matters what the details of the scam are.
Are you saying everyone who gets a kidney transplant because they lost all their kidneys actually dies and they just say it was something else?
LikeLike
Just as people think they know how to deal with so-called viruses because they believe in the statements of so-called scientists in the field of virology, immunology and molecular biology, so people think they know how to deal with transplants, because they believe, unconditionally, in the statements of transplant surgeons. That is why no one asks even the most basic questions to critically analyze the so-called allogeneic tissue and organ transplants. People do not wonder how it is possible for an allogeneic biological structure (tissue, organ) that has a different vibration than the transplant recipient’s body to be accepted by the transplant recipient’s body only because the transplant recipient is permanently poisoned with drugs considered to be immunosuppressants. That is why when they do an experimental transplant, doctors look for donors considered more compatible, ie who are as close as possible in terms of kinship to the recipient of organs. But who can say that, apart from the medical staff directly involved in the transplant process, they know the true success rate of experimental transplants of allogeneic tissues or organs? How many people would believe if they were told that after doctors had proven that allogeneic tissue and organ transplantation was not working they decided to go ahead with the scam called transplantology motivated by pride (so as not to be considered fools who play experimenting with people’s lives) and motivated by material interests? And how many people would think if they were told that apart from the cases where experiments are done, all transplantology is a medical staging in which surgeons mimic surgery (cutting the recipient’s tissues more or less deeply) so that patients and their relatives to think they had a transplant?
LikeLike
Nike, doctors believe in “viruses” and believe they are helping. Very few people in the industry understand that virology is a pseudoscience. The practitioners don’t have to be in on it. They just do great harm because they subscribe to a broken paradigm.
You are describing something entirely different: an intentional fraud by surgeons themselves covered up by all their support staff at hospitals around the world. Unless the number surgeons that ever do transplants is very very small, this gets into the fantastical. To get that many people doing wrong that deep in the system you need a broken belief system, not all of the many people who would have to be involved being in on it.
LikeLike
The level of forgeries and criminal scams in the field of Western Medicine is so high that people simply think it is crazy to believe this. Just as some of the great manipulators of the popular masses have confirmed: the more colossal a scam is, the more it is unconditionally believed.
LikeLike
The reality is that the fight to get rid of the criminal deceptions of allopathic medicine is not limited to the field of microscopic biology. The whole medicine that works on the principles of allopathy is a colossal scam.
LikeLike
I agree it’s not just microbiology, Nike, but most doctors are trying to help. The paradigm and incentives make it a scam in total but built of good intentions in most places. There are many corrupt doctors, too, but I don’t see how they’d ensure only corrupt ones (and only corrupt nurses, support staff, etc.) work in transplants.
The pattern in the rest of medicine is bad paradigms, presumptions, overspecialization, passing the buck, incentives, and bias confirmation creating a malign brew of death, with a peppering of corruption as well. That’s the kind of explanation I’m looking for. Conspiracy theory only works at the center of a scam, not at the edges. I need to know why doctors *think* they’re doing good with transplants yet aren’t, just like oncologists think they’re doing good with chemo yet aren’t and pediatricians think they’re doing good with vaccines yet aren’t. Explaining some of it with special examples of direct fraud is fine as well, but other than some people at the top the lion’s share would need to be explained by the mundane evil of people “doing the best they can” to be convincing.
LikeLike
A small part of the medical staff in Western Medicine is made up of criminal crooks, concerned with preserving their leadership positions, extremely high material privileges and special social glory.
Most of the medical staff in Western Medicine is made up of comfortable people, who refuse to think and who only blindly apply some protocols, concerned only with keeping their jobs, positions, salaries and social status.
Only an EXTREMELY small number of individuals in the medical staff of the Western Medical System critically analyze modern medicine based on the principle of anti-symptomatology that uses toxic pharmaceutical drugs and vaccines and all sorts of surgical, investigative, therapeutic and substitution procedures that are harmful.
Of course, the Western Medical System, in collaboration with the financial class, the political class, the repressive forces of the Police, the Prosecutor’s Office and the Justice and the Mass Media, do everything possible to ridicule and silence, by any means, those extremely few medical personnel that expose the colossal criminal character of the whole of Western Medicine.
LikeLike
I agree with all that, Nike. The whole system from work life to “education” to agriculture to diet to sickcare is culturally enforced and plenty of special interests capitalize on that.
LikeLike
PLEASE HELP!
I am currently arguing online with a virologist about ‘control experiments’. I am saying that his ‘control experiments’ are not scientifically valid.
He has has just sent me this picture with caption:
https://ibb.co/169fSNV
And he has also sent me this text:
“Our control experiments are perfectly valid. Any well designed study will include a mock inoculation along with serial dilutions as a negative control, with quantitative and/or qualitative differences in cpe.”
How am I suppose to respond to that? Has he got me or not?
LikeLike
Proper controls would be using the same fluids from healthy subjects, fluids from those said to have a similar disease like influenza, fluids from a person with lung cancer, testing each of the culture substances used alone and in combination (antibiotics, fetal bovine serum, antifungals, mediums) in varying concentrations, etc. In other words, controls should extend far beyond just mock infections which is what the image shows and all that they ever do.
LikeLike
@Mike Stone. First of all, thank you for replying. I am very grateful for your help. This is the situation now. I have replied to the virologist. He has replied back and said two things. The first thing he said was that I was being ridiculous. And the second thing he said was that the ONLY POSSIBLE REASON for the cpe must be that the infected sample contained a virus. Because the mock infection, which DID NOT contain a virus, DID NOT produce any cpe. Can you please help me formulate a reply to that?
LikeLike
Of course! I’m happy to help. I would ask him how he knows that it is not the added cell-toxic chemicals and/or foreign genetic material combined that is causing the observed CPE if they only do mock infections without fluids that do not contain the assumed “virus.” The only way to know for sure would be to do controls using fluids from healthy hosts and those suffering from similar symptoms of disease and treating the cultures exactly the same as those with the assumed “virus.” Doing the same thing without the fluids (i.e. mock infections) is not a proper control.
These articles may help as there are many known factors that cause CPE other than “viruses:”
https://viroliegy.com/2021/08/29/the-cytopathic-effect/
https://viroliegy.com/2021/09/04/creating-the-cytopathic-effect/
LikeLike
@ Mike stone. Thank you. That is exactly what I’m looking for. I’m sure even the brainwashed virologist I am talking to will understand that. I will let you know what he says next. Thanks again for your help.
LikeLiked by 1 person
Of course! I am excited to hear the response! 😉
LikeLike
@ Mike stone. My virologist has not replied today. Maybe I have struck a nerve or something. lol. However, if he does reply, I suspect he will try to bamboozle me be by going on about new age sequencing or metagenomic sequencing or something like that. So I think it would be wise to arm myself with a working knowledge of Stefan Lanka’s experiments where he managed to create an entire covid genome from yeast or something similar. Is there a sort of ‘idiots guide to Stefan Lanka’s experiments’ that I could read? The simpler the better, so I have a decent chance of understanding it. I have watched several of Stefan Lanka’s videos, and he is a genius, but he has a terrible German accent which I find difficult to understand. All suggestions are most welcome. Thanks again for helping me, Mike. I really appreciate it.
LikeLike
Hi Mike. Unfortunately, I don’t really know if there is a simple guide to Dr. Labka’s genomics data as I do not believe the full details of those experiments have been revealed yet. The best information I could find I put into this article:
https://viroliegy.com/2022/08/16/the-path-paved-by-dr-lanka/
In all honesty, if the virologist does try to bring up genomics, tell him that you will not discuss it as genomics is a red herring. I would stick to the lack of evidence of any particles claimed to be “viruses” being purified and isolated directly from the fluids of a sick host. As there is no valid independent variable, the scientific method has never been followed regarding virology. Thus, virology is by definition pseudoscience. Unless he can show you properly purified and isolated particles found in the fluids of humans that are proven pathogenic in a natural way, he’s got absolutely nothing. Genomics can not cover that up.
LikeLike
@Mike Stone and also @ClariFire. It looks like my virologist is not going to reply to me any more. I sent him a message explaining exactly why his ‘mock infections’ were not scientifically valid. And then I sent him another message further explaining that his methods do not have a valid independent variable. I have not heard anything back since. Anyway, Mike, thanks for helping me. Hopefully we have given this virologist something to think about.
P.S. I’m really pleased that the virologists claims can be completely refuted without having to delve into genomics and whatnot. Otherwise I would have been completely out of my depth. You guys are my heroes. Keep up the excellent work.
LikeLiked by 1 person
That’s too bad. I was hoping the virologist would stick around. If they do ever contact you again, ask if they would like to have a cordial conversation with me. I’d be more than happy to engage. Thanks for the update! 🙂
LikeLike
They tend to stop replying when they run out of playbook material, when they reach an area they haven’t thought about. They’ve almost invariably not thought far off the reservation.
Mock infections provide the necessary trappings of science, the window dressing that feels like control experiments – good enough to keep for a movie that has sciency stuff, good enough to maintain the suspension of disbelief in people who want to believe – while failing to cover the necessary ground scientifically. Much like voting makes us FEEL like we’re no longer living in an aristocracy, but I digress…
LikeLiked by 1 person
@Mike Stone. Hi Mike. I have news. Just when I thought my virologist was not going to reply to me any more, he goes and replies to me. But, unfortunately, it’s not that interesting. Instead of addressing the stuff I sent him about valid control experiments and independent variables etc that you told me about, he sent he a message saying that I need to ‘educate myself’. And he also sent me two links. This one about the Baileys:
https://dailysceptic.org/2022/10/18/will-the-virus-sceptics-ever-accept-the-evidence-that-proves-them-wrong/
And this video by Dr Wilson, a Ph.D molecular biologist, called “Virus deniers: the Flat Earthers of biology” which is here:
If this virologist is not willing to address the actual issues I sent him, then I think we might wasting our time with this particular individual. As you requested, though, I have asked him if he would be willing to have a cordial chat with you.
This is my new hobby now. Writing to virologists and trying to get them to admit that their whole enterprise is pseudo-science. I am retired and I have plenty of time on my hands.
I am also determined to gain enough knowledge to debunk every single point that Dr Wilson makes in his “Virus deniers: the Flat Earthers of biology” video. So, if you could give me a few links to read on that subject, that would be great.
I will let you know if I get any more virologists ‘on the hook’, as it will obviously be better if you talk to them instead of me.
All the best to you, my friend.
Mike.
LikeLiked by 1 person
Hi Mike! Sadly, they usually resort to ad hominem attacks and avoiding addressing anything when they have no legs left to stand on in the conversation. We see it all the time. You are definitely correct that further communication will most likely result in a waste of time. I am willing to talk to him (or future virologists) but when the personal attacks and logical fallacies come out, it is a sign that there is no more room left for discussion. Keep up the great work pushing them on the issue and educating yourself in the process! Speaking with these people was one of the best ways I learned as it forced me to research any claims they made or methods they threw out that I was unfamiliar with. I’m also happy to help in any way I can going forward. 🙂
LikeLike
@Mike Stone. Thank you for your kind offer of help, Mike. If I get another virologist ‘on the hook’, I will definitely be taking you up on it. By the way, I wondered if you might be interested in reading this:
https://www.jeremyrhammond.com/2022/10/17/answering-tom-cowans-five-simple-questions-for-virologists/?utm_source=substack&utm_medium=email
I tried to read it, but some of it was a bit complicated for me at the moment. I will understand it eventually, though. All the best, my friend. Goodbye for now.
Mike.
LikeLike
Hi Mike,
I have a friend who has been following our conversation and she has wanted to comment but has been having technical difficulties. She has been heavily involved in uncovering this hoax with myself and others. Her name is Carolyn Gutman Dey and she would love to help out in some way in contacting virologists/microbiologists in order to get some answers from them. If you’d like some help in your quest, she said to shoot her an email:
Forcgd@yahoo.com
LikeLike
Hi Mike,
I’m back. Someone on a motorcycle bumped into a ladder I was on. It knocked me off and I had to spend almost a week in hospital.
Anyway, when I am feeling a bit better, I will shoot your friend Carolyn an email, and see if we can come up with a plan.
Just wanted to let you know I am still fighting the good fight.
Mike.
LikeLiked by 1 person
I’m sorry to hear that you got injured but I’m happy you are on the mend. 🙂
LikeLike
When the virologist said, “You need to educate yourself,” I bet he meant, “I need to refresh myself on basic virology because I don’t have an answer to your question but it must be in there somewhere. And here, this other guy said you’re wrong so go argue with him.”
LikeLiked by 1 person
Absolutely! There is so much hidden in what they say. 😉
LikeLike
Mike, if one searches on this website (viroliegy.com) for the word “Visser”, no results are seen. May I suggest you write a post which includes that word in the title of any new article? By ‘Visser’, I refer to a Frank Visser, and in case you (and the other folks in the “no-virus” team = please alert them too!) are not already aware, – he has written several articles critiquing the “no-virus” position on his website – https://www.integralworld.net/readingroom.html
At that web page, scroll down and check out the many articles under the headings “VOLUME ONE: ONLINE CHAPTERS AND KINDLE EBOOK” and “VOLUME TWO: EXTRA ONLINE CHAPTERS”.
Besides those, there are some other relevant articles too on that web page, which have a bearing on the “virus” debate (search, for example, the terms “virus”, “covid”, “corona”; and also see the section ‘Most recent additions to this Reading Room’)
While a rebuttal of each of those articles may not be feasible, after reviewing every one of them, a general response that offers an overall rebuttal of the notable points therein (including the criticism of Dr. Lanka) may be considered. I suspect that there might be some back-and-forth once you set the ball rolling!
As always, keep up the good work!
LikeLike